köztes* intron a géneknek a képezők közé iktatott, fehérjéket nem kódoló, változó hosszúságú, sajátos nukleotidokkal kezdődő (GUA) és végződő (CAG) bázissora, amely ugyanolyan nukleotidokból áll, mint a képezők, csak bázissorrendjük más. (→génszerkezet, képezők) A köztesek hosszúsága változó (a legkisebbek <1000 bázispár, a legnagyobbak >10 kb), száma általában eggyel kevesebb a képezőkénél, de vannak bonyolultabb szerkezetek is, például az 5’-vég első köztese darabolva a továbbiakhoz kapcsolódik. A gének ~90%-ban fordulnak elő, összességében általában hosszabbak a képezőknél. A köztest tartalmazó géneket közteses* (discontinuous, split gene) nevezzük, szemben az egysejtűek folyamatos génjeivel. Néhány köztes fontos jeleket hordoz, de vannak gént kódolók is (nested genes). A közteseknek négy törzsökös elemét különböztetjük meg:
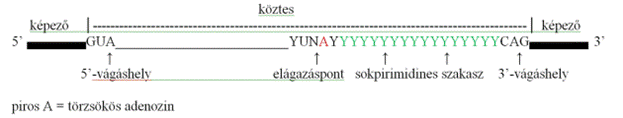 • Az 5’-végi vágáshelyet* (5’ splice site – 5’SS);
nevezik adó (donor) vágáshelynek is. az 5’-végi képező itt válik el a
köztestől. GU-val kezdődik.
• Az 5’-végi vágáshelyet* (5’ splice site – 5’SS);
nevezik adó (donor) vágáshelynek is. az 5’-végi képező itt válik el a
köztestől. GU-val kezdődik.
• A 3’-végi vágáshelyet* (3’splice site – 3’SS); nevezik kapó (acceptor) vágáshelynek is. AG-vel végződik.
• A sok pirimidint (Y jelöli) tartalmazó szakaszt a 3’-vágáshelyhez csatlakozva (sokpirimides szakasz*, polypyrimidine tract, PPT).
• Az elágazáspontot* (branchpoint, branch point sequence, BPS), amelyben törzsökös adenozin (piros A) van. Ez a hurok forma kialakításáért felelős.
A köztes bázisainak nincs szerepe a fehérjeképződésben, ezért a gének „értelmetlen” (nonsense) szakaszának is nevezik, szemben a képezővel, amely a gén „értelmes” (sense) szakasza, mivel ez kódolja a polipeptidet, határozza meg a gén kifejeződését.
Feltételezik, hogy a köztesek már megvoltak a fejlődés kezdetén, de helyhiány miatt kiszorultak az egysejtűek génjeiből.
Az 5’ és a 3’-vágáshely csorbái megzavarják a fehérjék képződését; számos örökletes betegség okozói.
A köztesek és képezők együttese teszi lehetővé a molekulafejlődést, új gének létrejöttét a testvér kromoszómák képezőinek az átrendeződés alatti cserélődéseivel. (→génszerkezet, képező)