G-fehérje-kapcsolt jelfogó G
protein-coupled receptor, GPCR (egyéb
nemzetközi nevei: seven-(pass)-transmembrane domain receptor, 7TM receptors, 7 membrane spanning, heptahelical
receptor, serpentine receptor, G protein-linked receptors, GPLR). A hazai irodalomban használatos a 7TM-jelfogó (7 transmembrane
[7TM] domain) elnevezés is.
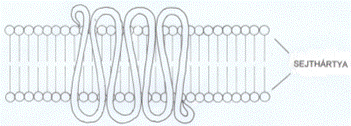 A 7TM-fehérjék a külső és belső felszínen hurkokkal
összekapcsolt hét sejthártyán áthaladó α-csavarulattal rögzülnek a
sejthártyában; ezen kívül a polipeptid lánc mindkét vége is hurkot formál - az
N-vég a sejten kívül, míg a C-vég a sejten belül található. A sejthártyán
kívüli rész köti a jelvivőt, a sejtbe nyúló hurkok közül kettő pedig a
háromalegységes G-fehérje α-alegységét; ez utóbbi esemény általában akkor
következik be, amikor a jelfogó gerjesztődik a jelvivő hatására, de az is
előfordul, hogy a G-fehérje a jelfogó tevőtlen állapotában is hozzá kapcsolt..
(Az ábrát Teleki Katalin készítette.)
A 7TM-fehérjék a külső és belső felszínen hurkokkal
összekapcsolt hét sejthártyán áthaladó α-csavarulattal rögzülnek a
sejthártyában; ezen kívül a polipeptid lánc mindkét vége is hurkot formál - az
N-vég a sejten kívül, míg a C-vég a sejten belül található. A sejthártyán
kívüli rész köti a jelvivőt, a sejtbe nyúló hurkok közül kettő pedig a
háromalegységes G-fehérje α-alegységét; ez utóbbi esemény általában akkor
következik be, amikor a jelfogó gerjesztődik a jelvivő hatására, de az is
előfordul, hogy a G-fehérje a jelfogó tevőtlen állapotában is hozzá kapcsolt..
(Az ábrát Teleki Katalin készítette.)
Ez a legnagyobb jelfogócsalád, emberben 800-nál több
génváltozatot ismerünk. Az általuk kötött jelvivők szintén igen változatosak: a
fotontól (rodopszin) az ionokon (H+, Ca++), a kis
molekulákon (odor- [szaglás-] molekulák) és a lipideken át a viszonylag nagy
polipeptidekig (FSH, LH, trombin, WNT) terjed. A 7TM-jelfogókat öt alcsaládba
(rodopszin-, szekretin-, glutamát-, adhéziós és a Frizzled típusú receptorok)
sorolják. Az általuk indított jelközvetítések egymásba is kapcsolódnak,
behálózzák az egész sejtet.
G-fehérje-kapcsolt jelfogó kináz G-protein-coupled receptor kinase, GRK szerin-treonin-kináz. Hét fajtája ismert; ezeket három
alcsaládba sorolják hasonlóság alapján (GRK1 [rodopszin-kináz] és GRK7; GRK2 és
GRK3; valamint GRK4–6). Három gomolyuk van: középen a sarkalló (foszforilező),
mely a PKA és PKC-hez (fehérjekináz-A, -C) hasonló szerkezetű; a szabályozó
gomoly az N-végen és a változó gomoly a C-végen. A GRK-k a G-fehérje-kapcsolt
jelfogók sajátos foszforilezése mellett sok más fehérjét is foszforileznek
(→epidermal growth factor [EGF], platelet-derived growth factor [PDGF] jelfogó, tubulin, R-SMAD stb.). A GRK-k által kapcsolt foszfátcsoport gátló hatású a
G-fehérje-kapcsolt jelfogóra; szerepük elsősorban a jelzésberekesztésben van.
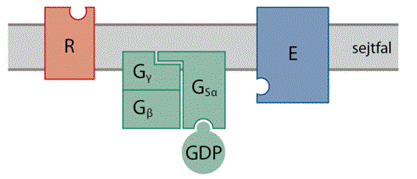 G-fehérje-kapcsolt jelközvetítés A
jelfogó sejtkívüli vájolatához (a vázlatos ábrán R jelöli) kapcsolódik a
jelvivő, és megváltozik a jelfogó szerkezete, aminek következtében a sejtbeli
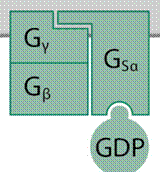
végen kötőhely válik szabaddá. Ehhez kapcsolódik a háromegységes (Gα, Gβ és Gγ)
G-fehérje α-alegysége, rendszerint egyféle, de – a jelfogó természetétől
függően – társulhat többféle is. A kötődés következtében módosul a Gα
szerkezete, megnyílik a GDP-t kötő zseb, és leválik a GDP. A sejtben a GTP
tízszer nagyobb töménységben van jelen, és – a harmadik foszfát miatt – nagyobb
hajlammal kötődik, mint a GDP, ezért a levált GDP helyét azonnal egy GTP
foglalja el. A GTP társulására megváltozik a teljes háromalegységes G-fehérje
szerkezete; a GTP-t kötő Gα tevősödik, egyrészt elválik a Gβ–Gγ-tól, másrészt
szerkezetváltozása következtében képes társulni a célenzimmel és tevősíti. A
célenzim rendszerint sejthártyabeli enzim (az ábrán E jelöli). A Gα GTPáz
tulajdonsága miatt egy idő után vízbelépéssel bontja a GTP-t GDP-re és foszfátra..
A GDP-t kötő Gα ismét tevőtlen állapotba kerül, újra egyesül és Gβ–Gγ-val, és a
folyamat kezdődhet elölről.
G-fehérje-kapcsolt jelközvetítés A
jelfogó sejtkívüli vájolatához (a vázlatos ábrán R jelöli) kapcsolódik a
jelvivő, és megváltozik a jelfogó szerkezete, aminek következtében a sejtbeli
végen kötőhely válik szabaddá. Ehhez kapcsolódik a háromegységes (Gα, Gβ és Gγ)
G-fehérje α-alegysége, rendszerint egyféle, de – a jelfogó természetétől
függően – társulhat többféle is. A kötődés következtében módosul a Gα
szerkezete, megnyílik a GDP-t kötő zseb, és leválik a GDP. A sejtben a GTP
tízszer nagyobb töménységben van jelen, és – a harmadik foszfát miatt – nagyobb
hajlammal kötődik, mint a GDP, ezért a levált GDP helyét azonnal egy GTP
foglalja el. A GTP társulására megváltozik a teljes háromalegységes G-fehérje
szerkezete; a GTP-t kötő Gα tevősödik, egyrészt elválik a Gβ–Gγ-tól, másrészt
szerkezetváltozása következtében képes társulni a célenzimmel és tevősíti. A
célenzim rendszerint sejthártyabeli enzim (az ábrán E jelöli). A Gα GTPáz
tulajdonsága miatt egy idő után vízbelépéssel bontja a GTP-t GDP-re és foszfátra..
A GDP-t kötő Gα ismét tevőtlen állapotba kerül, újra egyesül és Gβ–Gγ-val, és a
folyamat kezdődhet elölről.
Sokféle enzim válik működővé a GTP-kötött Gα-fehérjével
kapcsolódva. Ilyen enzimek többek között az adenilát-cikláz, amely másodlagos
hírvivőt, cAMP-t állít elő ATP-ből; a foszfolipáz-Cβ (PLCβ), amely két
másodlagos hírvivőt, diacilglicerolt és inozitol-trifoszfátot is létrehoz; a
cGMP-t bontó cGMP-foszfodiészteráz stb. Ismerünk G-fehérjékkel szabályozódó,
ún. G-fehérje-vezérelt ioncsatornákat; köztük vannak Cl-, K-, Ca- és
Na-csatornák is. Összességében igen sokféle folyamat (citokinek, hormonok
képződése, ingerület-átvezetés, izomösszehúzódás, sejtmozgások, szervfejlődés,
tanulás, emlékezés stb.) szabályozódik a G-fehérje-kapcsolt jelfogók általi
jelközvetítéssel; ezek kapcsolódnak egymással is, mintegy hálózatot képeznek;
elengedhetetlenek a szervezet egyensúlyállapotának megtartásában.
A jelfogónak a G-fehérjét serkentő hatása kétféleképpen
szűnhet meg:
• Azonnal, ha leválik a jelvivő.
• A GRK (G-protein-coupled receptor kinase)
hatására; ez a szabályozás fő formája. A GRK sajátosan csak a serkentett
jelfogót foszforilezi, a jelfogón kialkult foszfátmintázatot felismerve
hozzáfűződik a β-arrestin (β1/2-arrestin) fehérje, és fizikailag megakadályozza
a jelfogó–G-fehérje társulást. Megszűnik a jelátadás, és megkezdődik a
jelfogó–jelvivő bekebelezése: a β-arrestin hozzákapcsolja a jelfogót a
bekebelezést végző rendszerhez (→AP2 [adaptin], klatrin). A jelfogó a
jelvivővel együtt, hólyagocskába zárva, bekerül a sejtbe. A beltestecsben
(endosome) elválasztódnak az újrahasznosuló jelfogók a lebontandóktól; az
előbbiek ismét a sejthártyába kerülnek, az utóbbiak az emésztőtestecsbe
(lysosome), ahol teljesen lebomlanak.