CG- (citozin, guanin) mintázatok CG-rich secondary structures másodlagos DNS-szerkezetek, a DNS CG-gazdag részeiben, mint a CpG-szigetek, alakulnak ki. Kétféle CG-mintázat, a guaninnégyes (G-négyes) és az i-mintázat ismert; ezek elsősorban a gének 5’-végén, az átíródás kezdetétől felfelé, a génátíródást serkentő részeken (transcription-enhancing nucleosome-depleted promoter regions), továbbá az 5’-UTR-ben és a kromoszómavégeken (telomeric regions) találhatók.
guanin guanine kettős kötéssel társított két pirimidin-imidazol gyűrűt tartalmazó molekula (C5H5N5O), purinszármazék. Hatáscsoportja az NH2 (aminocsoport). Egyike a DNS-ben és az RNS-ben lévő négy alapvető bázisnak. (→guaninnégyes)
Találatok címszavakban (4 szócikk):
GEF ( guanine exchange factor) cserélőfehérje a GDP–GTP cserélődést hozza létre: a GEF kötődik valamelyik GTPáz-hoz (pl. RAB, RHO fehérje), ennek következtében változik a GTPáz térszerkezete, és leválik a GDP. A szabadon maradt kötőhelyhez a plazmában bőven lévő GTP kötődik. Pl. a G-feréjével kapcsolt jelátadásnál a 7TM-jelfogó a GEF: a G-fehérje kapcsolásával (szerkezetváltozás) segíti elő a GDP leválását.
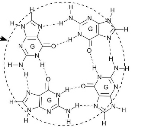 G-négyes (guaninnégyes)* DNA G-quadruplex (GQ) structure, G-quartet egymásra
tevődött négytagú guanintársulásokból alakult állékony másodlagos DNS-szerkezet.
G-négyes (guaninnégyes)* DNA G-quadruplex (GQ) structure, G-quartet egymásra
tevődött négytagú guanintársulásokból alakult állékony másodlagos DNS-szerkezet.
▪ Négytagú guanintársulás négy egymást követő guanin kapcsolódásából jön létre. A guaninok Hoogsteen-féle hidrogénkötésekkel (Hoogsteen hydrogen-bonded guanines, G-quartets) kötődnek egymáshoz. Azaz – ellentétben a Watson–Crick-féle bázispárosodással, amelyben a hidrogénhidak az adenin és a timin, illetve a guanin és citozin között jönnek létre – a négytagú guanintársulásokban a hidrogénhidak a guaninok között alakulnak ki.
 Két-három ilyen négytagú guanintársulás tevődik egymás
fölé, aminek következtében a DNS többszörösen elhajlik, visszafordul,
párhuzamos szálakat kialakítva. Az ábrán három négytagú guanintársulás egymásra
tevődéséből alakult G-négyes látható. Állékony másodlagos szerkezetek, a
guaninban gazdag egyszálú DNS-ben keletkeznek, rögzítik az egyszálú DNS-t. Pl.
a végrészek túlnyúlásaiban. Gyakoriak a CpG szigetekben. (→végrész)
Két-három ilyen négytagú guanintársulás tevődik egymás
fölé, aminek következtében a DNS többszörösen elhajlik, visszafordul,
párhuzamos szálakat kialakítva. Az ábrán három négytagú guanintársulás egymásra
tevődéséből alakult G-négyes látható. Állékony másodlagos szerkezetek, a
guaninban gazdag egyszálú DNS-ben keletkeznek, rögzítik az egyszálú DNS-t. Pl.
a végrészek túlnyúlásaiban. Gyakoriak a CpG szigetekben. (→végrész)
Számos fehérje – kivált az átírásfehérjék, továbbá helikázok, kromatinmódosítók, állapotszabályozók – ezt ismerik fel, ehhez kapcsolódnak.
A G-négyesek a hozzájuk kapcsolódó átírásfehérjékkel együtt a génátíródás „ki-bekapcsolói”; a fehérjékkel kapcsolódva sokszorozzák az átíródást. Feltételezik, hogy a rákos átalakulásban is szerepük van: a törzsbázissorban vagy a környékén lévő G-négyesek ugyanis jelentősen fokozhatják a daganatserkentő gének (pl. cMYC) tevékenységét.
guanine nucleotide-binding proteins →G-fehérjék
guanin nukleotidcserélő tényező guanine nucleotide exchange factor (→nukleotidcserélő tényező)
Találatok alcímszavakban (1 szócikk):
nukleotid nucleotide bázisból (purin-/pirimidinbázis), szénhidrátból (ribóz/dezoxiribóz) és a szénhidráthoz kapcsolódó foszfátból álló molekula, azaz a nukleozidok foszfátészterei, nukleozid-foszfátok. A nukleozidoktól tehát csupán a foszfátcsoporttal különböznek. A foszfátok kapcsolódása szerint a ribonukleozidból háromféle – 2’-, 3’- és 5’-foszfát –, a dezoxiribonukleozidból pedig kétféle – 3’- és 5’-foszfát – azonmás vezethető le. A számok a ribóznak, dezoxiribóznak azokat a szénatomjait jelölik, amelyekhez a foszfátcsoport kapcsolódik (→adenozin).
Egy nukleozidhoz több foszfát is kötődhet: két foszfát kapcsolódásakor nukleozid-difoszfát, három kötődésével nukleozid-trifoszfát keletkezik. A foszfátok egymáshoz, és nem külön szénatomhoz kapcsolódnak, nagyenergiájú (savanhidril) kötésekkel. Az egymáshoz kapcsolódó foszfátokat α, β és γ jelzéssel különböztetjük meg. Az α jelzésű foszfát kapcsolódik észterkötéssel a szénhidráthoz; a másik kettő foszfohidridkötéssel egymáshoz; pl. az adenozinból két foszfát kapcsolódásakor adenozin-difoszfát (ADP), három kötődésekor adenozin-trifoszfát (ATP) képződik (→adenozin). A három foszfátot tartalmazó molekulák (trifoszfátok) a sejtek legjelentősebb energiahordozói. Az energiát a második és harmadik foszfátkötésben tárolják. Energia leadásakor az utolsó (γ-helyzetű) foszfát hasad le vízzel bontással, és energia szabadul fel; ez a sejtek működésének az alapvető energiaforrása.
A nukleotidok elnevezése a bázisból vezethető le. Például az adeninből keletkező nukleozid az adenozin, amelynek foszfát származéka, vagyis a nukleotid, az adenozin-foszfát – mivel egyetlen foszfátot tartalmaz, adenozin-monofoszfát (AMP). A két foszfátot tartalmazó az adenozin-difoszfát (ADP), a hármat tartalmazó az adenozin-trifoszfát (ATP) (→adenozin). Hasonlóan: guanozin-trifoszfát, timidin-trifoszfát (TTP), citidin-foszfát.
A nukleozid-monofoszfát só/észter származékait -ilát végződéssel jelöljük. Pl.:
adenozin-foszfát (AMP) → adenilát; dezoxiadenozin-foszfát (dAMP) → dezoxiadenilát
guanozin-foszfát (GMP) → guanilát; dezoxiguanozin-foszfát (dGMP) → dezoxiguanilát
citidin-foszfát (CMP) → citidilát; dezoxicitidin-foszfát (dCMP) → dezoxicitidilát
timidin-foszfát (TMP) → timidilát
uridin-foszfát (UMP) → uridilát
Ezekkel az elnevezésekkel szokásosan az enzimek neveiben találkozunk (adenilát-kináz, timidilát-szintetáz stb.).
A nukleotidok száma szerint megkülönböztetünk egy nukleotidból álló vegyületet, amelynek a nemzetközi neve: mononucleotide. Az oligonucleotide rövid, néhány nukleotidból álló egyszálú DNS-/RNS-töredék. A sok nukleotidból képződő vegyület pedig a polynucleotide (nukleinsav).
nukleotidcserélő tényező (guanin-nukleotid cserélő tényező) nucleotide exchange factor enzim, a guanin nukleotidkötő fehérjéhez kapcsolódott GTP-t cseréli GDP-re.
nukleotid-másolódószál* antisense oligonucleotide, ASO rövid, néhány nukleotidot tartalmazó, mesterségesen előállított másolódó egyszálú nukleotidsor. Számos felhasználási területe van: a gyógyszerkészítéstől az RNS-ek, fehérjék befolyásolásáig.
nukleotidmásulás* azonos a bázismáulás nevezettel. (→bázismásulás)
Részleges egyezések (4 szócikk):
adenin/guanin metilezés (többféle lehetséges) megváltoztatja a bázisok viselkedését. A metil-guanin ugyanis a citozin helyett a timinnel kötődik, a metil-adenin pedig nemcsak a timinnel, hanem a többi bázissal is. A metilcsoportot az adeninhez a DAM (dioxi-adenin-metiláz) kapcsolja.
G-fehérjék guanine nucleotide-binding proteins GTP-kötő fehérjék, számos jelközvetítést és megszámlálhatatlan sejtfolyamatot szabályoznak; miután a kötött GTP-t GDP-vé képesek alakítani, GTPázoknak is nevezik őket. Két formájuk ismert: a három (α, β és γ) alegységes és az egy alegységes fehérje; az előbbieket nevezik nagynak, az utóbbiakat kicsinek is.A három alegységes G-fehérjék esetében valójában csak az α alegységek rendelkeznek a GTP-kötés képességével, ezek többfélék lehetnek: a leggyakoribbak a Gαs (serkentő) és a Gαi (inhibitor, gátló) G-fehérjék, de van Gαq- és Gα12-fehérje is.
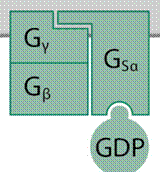 nagy G-fehérje csak az α-alegysége (Gα) képes a GTP kötésére (a
kapcsolódás helye a GTP-kötő zseb). Az α-alegységhez kapcsolódó β- és
γ-alegységet (Gβ és Gγ) – mivel nem kötnek GTP-t – nem is sorolják a G-fehérjék
családjába. Miután az α- és a γ-alegységhez elektron-kötéssel lipid (zsírsav)
molekula is kapcsolódik (lipid-horgony), ezek a sejthártya belső rétegébe
illeszkednek, míg a β-alegység a γ-val társulva vesz részt a szerkezet
kialakításában. A Gβ WD40 ismétleteket, gomolyt tratlmaz, állványfehérjeként
lehetővé tesz fehérjekölcsönhatásokat. A hármas csak akkor marad együtt, ha az
α-alegység GDP-kötött, gerjesztetlen állapotban van. A GTP-vel kapcsolt
α-alegység viszont elválik a másik kettőtől: így fejti ki a hatását. A GTP-kötött
α-alegység leválása után szabaddá vált βγ-alegység is képes lesz más
célfehérjékhez kapcsolódva azokat gerjeszteni. Az α-, a β- és a γ-alegységeket
sokféle különböző gén kódolja, ezért számos változatuk van: társulásukkal több
ezerféle háromalegységes G-fehérje keletkezhet; mind részt vehet más-más
jelátvitelben. Az alegységek a sejthártyáhozhoz kapcsolódva egyesülnek
háromalegységes G-fehérjévé. (Az ábrán a szürke rész a sejthártyát jelöli. Gsα
= serkentő formájú α-alegység. (→G-fehérje-kapcsolt jelfogó, G-fehérjés jelközvetítés)
nagy G-fehérje csak az α-alegysége (Gα) képes a GTP kötésére (a
kapcsolódás helye a GTP-kötő zseb). Az α-alegységhez kapcsolódó β- és
γ-alegységet (Gβ és Gγ) – mivel nem kötnek GTP-t – nem is sorolják a G-fehérjék
családjába. Miután az α- és a γ-alegységhez elektron-kötéssel lipid (zsírsav)
molekula is kapcsolódik (lipid-horgony), ezek a sejthártya belső rétegébe
illeszkednek, míg a β-alegység a γ-val társulva vesz részt a szerkezet
kialakításában. A Gβ WD40 ismétleteket, gomolyt tratlmaz, állványfehérjeként
lehetővé tesz fehérjekölcsönhatásokat. A hármas csak akkor marad együtt, ha az
α-alegység GDP-kötött, gerjesztetlen állapotban van. A GTP-vel kapcsolt
α-alegység viszont elválik a másik kettőtől: így fejti ki a hatását. A GTP-kötött
α-alegység leválása után szabaddá vált βγ-alegység is képes lesz más
célfehérjékhez kapcsolódva azokat gerjeszteni. Az α-, a β- és a γ-alegységeket
sokféle különböző gén kódolja, ezért számos változatuk van: társulásukkal több
ezerféle háromalegységes G-fehérje keletkezhet; mind részt vehet más-más
jelátvitelben. Az alegységek a sejthártyáhozhoz kapcsolódva egyesülnek
háromalegységes G-fehérjévé. (Az ábrán a szürke rész a sejthártyát jelöli. Gsα
= serkentő formájú α-alegység. (→G-fehérje-kapcsolt jelfogó, G-fehérjés jelközvetítés)
kis G-fehérje 180–300 aminosavat tartalmaz; egyetlen egység alkotja. A kis G-fehérjék legelőször felfedezett tagja a RAS fehérje, amely lipid-horgonnyal a sejthártya belső felszínén rögzül (a háromalegységes G-fehérjék α-alegységéhez hasonlóan). Ugyanakkor a kis G-fehérjék közé tartoznak a sejtplazmában található oldott fehérjék is. A kis G-fehérjék a sejtműködés szinte minden folyamatában közreműködnek.
G-quadruplex →guaninnégyes
8-oxoG (oxoguanin nyolcas, 8-oxoguanine) a guanin elektronleadásából keletkező átrendeződése a guaninban gazdag ismétletekben, például a végrészek túlnyúlásaiban. Másulás következménye.