 kromatin chromatin a DNS-ből és a körülötte szerveződő hisztonokból
és más fehérjékből áll. Alapszerkezetét egy összességében negatív töltésű 11
nm-es szál képezi, amelyben a magtestecs (nucleosome) és az összekötő DNS-szál
~200 bázispárnyi mintázatai ismétlődnek. Másként: a kromatin fehérjékbe
csomagolt DNS. Mintegy 30 millió magtestecs keletkezik a sejtmagban. A kromatin
a magállomány zömét adja. Fehérjéi tömörítik, feltekercselik és a nem tevékeny
kromoszómarészeken – átmenetileg vagy véglegesen – teljesen becsomagolják a
DNS-t; elzárva a környezetétől. Ez teszi lehetővé, hogy a testi sejtjeinkben
~1,8 m hosszú DNS elférjen a néhány mikrométeres magban. A kromatin továbbá nagyon
fontos szabályozó a DNS átíródásában is, mert meghatározza, milyen mértékben
képesek az átírásfehérjék, az RNS polimeráz illetve más, a folyamathoz
szükséges fehérjék a DNS-hez hozzáférni. (Az ábrán a kék korongok a
magtestecsek, a DNS köréjük tekeredik – készítette Baghy Kornélia)
(→magtestecs)
kromatin chromatin a DNS-ből és a körülötte szerveződő hisztonokból
és más fehérjékből áll. Alapszerkezetét egy összességében negatív töltésű 11
nm-es szál képezi, amelyben a magtestecs (nucleosome) és az összekötő DNS-szál
~200 bázispárnyi mintázatai ismétlődnek. Másként: a kromatin fehérjékbe
csomagolt DNS. Mintegy 30 millió magtestecs keletkezik a sejtmagban. A kromatin
a magállomány zömét adja. Fehérjéi tömörítik, feltekercselik és a nem tevékeny
kromoszómarészeken – átmenetileg vagy véglegesen – teljesen becsomagolják a
DNS-t; elzárva a környezetétől. Ez teszi lehetővé, hogy a testi sejtjeinkben
~1,8 m hosszú DNS elférjen a néhány mikrométeres magban. A kromatin továbbá nagyon
fontos szabályozó a DNS átíródásában is, mert meghatározza, milyen mértékben
képesek az átírásfehérjék, az RNS polimeráz illetve más, a folyamathoz
szükséges fehérjék a DNS-hez hozzáférni. (Az ábrán a kék korongok a
magtestecsek, a DNS köréjük tekeredik – készítette Baghy Kornélia)
(→magtestecs)
Találatok címszavakban (13 szócikk):
eukromatin, heterokromatin (→kromatinformák)
kromatinállapot A kromatin kétféle állapotban, lazult (laza kromatin) és tömörödött formában van jelen a nyugalmi sejtmagban. Az utóbbinak is két formáját, az állandóan tömörödött (constitutive heterochromatin) és az időszakosan tömörödött (facultative heterochromatin) kromatint különböztetjük meg. A kettő között lényegi különbség van: az állandóan tömörödöttben nincsenek, az időszakosan tömörödöttekben vannak gének, csak némítva.
A lazult forma nemzetközi elnevezése euchromatin, a tömörödötté heterochromatin. Ezt a hazai nevezettár is átvette magyarosan írva: eukromatin és heterokromatin formában. A heterochromatin elnevezés megtévesztő: arra utal, hogy valami más molekula is van benne, noha nincs. Az eu- előtag jelentése ’jól, könnyen’; ellentéte a dys- előtag. Ennek tükrében az euchromatin a jó kromatin. Akkor a heterochromatin a rossz kromatin, ami nem igaz, hiszen igen jelentős szerepe van a sejt működésében. A fentiek miatt a laza kromatin és a tömörödött kromatin elnevezés kifejezőbb, megfelelőbb. Mivel az állandóan és időszakosan tömörödött kromatin között lényegi különbség van: célszerű külön névvel illetni őket. Az állandóan tömörített kromatinnak tömör kromatin*, az időszakosan tömörödöttnek tömörödő kromatin* lehet a neve.
■ A laza kromatin a tevékeny kromatin. Génszerkezeteket tartalmaz, és lehetővé teszi azok átíródását: a laza szerkezete következtében az átírásfehérjék, enzimek, RNS-ek hozzáférnek a DNS-hez. Jellegzetes a hisztonvégek (histone tails) acetilezett állapota. A magtestecsek egyvonalban sorakoznak, gyöngyfüzér alakzatban vannak; a köztük lévő távolság viszonylag nagy, miként az összekötő DNS is, több a hozzáférhető hely.
A laza kromatin a kromatinállomány ~90%-át teszi ki, főleg a sejtmag belsejében helyezkedik el. A tevékeny sejtmagokban sok a laza kromatin, és mivel helyigényes, hiszen kevésbé tömörített, a sejtmagok nagyobbak. A laza kromatin a sejtosztódás korai S-szakaszában másolódik.
■ Tömörödött kromatin a kromatin tétlen formája abban az értelemben, hogy nincs benne génátírás. A kétféle tömörödött kromatin, a tömör és a tömörödő kromatin szerkezete és jelentősége is eltérő.
▪ Tömör kromatin olyan kromoszómarész, amelyik minden sejtben jelen van, kizárólag tömörödött formában; ismétletekből és ugrálatokból tevődik össze, nem íródik át. A kromoszómák alapvető rögzítő szerkezeti eleme, a befűződések és a végrészek körüli DNS-t alkotja. Jellemzi a sokalakúság, és hogy C-sávot tartalmaz. Bizonyos sejtekben sajátos helyzetű. A nyugalmi sejtmagban összenyomott, sötéten festődő szemcsék formájában látható.
▪ Tömörödő kromatin olyan tömörödött kromatin, amelyikben nincs megnyilvánuló átíródás, de külső hatásokra átmenetileg átalakulhat laza kromatinná, és átíródhat. Jellemzően hosszú elszórt ugrálatokat tartalmaz, és vannak benne gének is, de nincs C-sáv. Meglehetősen egyforma, de változékony szerkezetű, nem sokalakú. A néma X-kromoszómát ez alkotja. Feltételezhető, hogy a sejtfejlődés folyamán azok a gének, amelyek az elődsejtekben tevékenyek, de az érett sejt működésében már szükségtelenek, tömörödő kromatin képződésével némulnak el.
A tömörödött kromatinban kevés az acetilezett hiszton, viszont sok a 9-es lizinen két-háromszorosan metilezett H3 (H3K9me2/3), amelyekhez rendre a tömörödött kromatinfehérjék, például a HP1 (heterochromatin protein 1) kapcsolódik, amely más fehérjéket toboroz, és hidat képezve a magtestecsek között, tömöríti őket. A tömörített kromatinban a magtestecsek szorosan vannak egymás mellett, rövid az összekötő DNS, és a magtestecssorok sokszorosan hajtódva helyezkednek el, egyre vastagabb szálakat képezve.
Ez a kromatin a késői S-szakaszban másolódik, a számcsökkentő sejtosztódásban nem rendeződik át. Sajátos tulajdonsága, hogy átterjed a szomszédos részekre is, ha a körülmények úgy hozzák. Például, ha a laza kromatin valamely tevékeny génje áthelyeződik a tömörödött kromatinba vagy közeli részbe, tétlenné válhat, nem íródik át. Ezt nevezik áttevődéses helyzethatásnak* (position effect variegation, PEV).
Lévén tömörített, a tömörödött kromatin kisebb helyen elfér, ezért azok a sejtmagok, amelyekben a kromatin zöme tömörödött, rendszerint kicsik, zsugorodottak (picnotic nuclei). A kevésbé zsugorodott magokban a tömörödött kromatin szemcsék formájában látható. A kromoszómák kromatinná visszaalakulásakor keletkezik („kromoszóma-maradványok”), vagyis a sejtosztódás származékai.
A laza és a tömörödött kromatin szerkezet alapvetően csak a magtestecsek számában tér el; az utóbbiban sokkal sűrűbben vannak. Mindkettő változékony szerkezet. Vegyi tulajdonságaik azonban eltérnek: a tömörödött kromatin erősebben festődik, több elektront tömörít (elektronmikrográfia), ellenállóbb a DNS-t oldó molekuláknak.
kromatinállapot szabályozása. Három eleme van: a hisztonmag-módosítás, a H1-változás és a DNS állapota.
■ A hisztonmag módosítása:
● Egyrészt hatócsoportoknak (acetilezés, metilezés stb.) a hisztonok N-végeihez (histone tail, hisztonfarok) való kapcsolódással valósul meg. Ez a hisztonmagot módosító enzimek (hisztonmódosítók) közreműködésével jön létre. A hisztonmaghoz sokféle hatócsoport kötődhet:
▪ A legjelentősebb a hisztanmag, főleg a H3 és H4 N-végi lizin oldalláncainak acetilezése és metilezése.
- A hisztonok N-végi acetilezése lazítja és tevősíti a kromatint, akadályozza a kromatin tömörödést. Az N-végi acetilezés amidkötés létrehozásával csökkenti a lizinek pozitív töltését, gyengül a hisztonmag és a negatív töltésű DNS közötti töltéskölcsönhatás, fellazul a szerkezet. Lehetővé válik, hogy az acetilezett hisztonhoz közeli génhez bromogomolyú átírásfehérjék kötődjenek. A bromogomolyt tartalmazó fehérjék elősegítik a kromatin-ATPázok kötődését a hisztonokhoz. Ezek az enzimek elcsúsztatják a magtestecseket, miközben a DNS egy része szabaddá válik, például az indító, és kötődhet hozzá az átírásfehérje.
A hisztonok acetilezését a hiszton-acetiltranszferáz (histone acetyltransferase, HAT, más néven histone acetylase, hiszton-acetiláz) végzi: a hiszton N-végi lizinjére acetilcsoportot visz át az acetil-koenzim-A-ról; ε-N-acetil lizin keletkezik. A hiszton-acetilázokat nevezik lizin-acetilázoknak is, mert mindig a lizinről távolítják el az acetilcsoportot, és nemcsak a hisztonok lizinjéről.
Az acetilezett hisztonok dezacetilezése ellenkező hatású; tömörödik a kromatin. Az acetilcsoport eltávolításával fokozódik a hiszton pozitív töltése, ezért szorosabban köti a DNS-t, tömöríti a kromatint. A dezacetilezést rendszerint metilezés követi. A dezacetilezést a hisztonok ε-N-acetil lizinjének acetilcsoportját eltávolító enzim, a hiszton-dezacetiláz (histone deacetylase, HDAC) sarkallja.
- A hisztonok N-végi metilezése ellentétes hatású, mint az acetilezése, például a H3 9-es lizinjének metilezése némítja a DNS-t, serkenti a kromatin tömörödését. Ugyanígy hat az N-végi arginin metilezése.
▪ Fontos az N-végi szerin/treonin oldalláncok foszforilezése, defoszforilezése. Például a H1 defoszforilezésével megnyúlik a kromatin. Ennek a sejtosztódásban van jelentősége: az S-szakaszban a H1 alig foszforilezett, lehetővé téve a másolódást; a G2–M átmenetnél viszont kifejezetten foszforilezett, a kromatin tömörödik.
● A hisztonmag módosítását jelentősen befolyásolják a töltésviszonyok: A hisztonmag végei erősen pozitívak a bennük lévő arginin és lizin miatt, lehetővé téve sokrétű kapcsolódást a DNS-sel, sőt távoli molekulákkal és a szomszédos magtestecsekkel is. A magtestecsek hisztonvégződéseinek összekapaszkodása a kromatintömörödés egyik alapfolyamata. Bizonyos nem hiszton fehérjék, például a kohezin, kifejezetten serkentik a magtestecsek társulását, vagyis a kromatin tömörödését.
A DNS szorosan kapcsolódik a hisztonmaghoz töltéskötésekkel, amelyek a pozitív töltésű aminosavak és a negatív töltésű DNS-váz foszfátcsoportjai között alakulnak ki.
A kromatin szerveződését tehát a magtestecsek kapcsolódásai, mozgása alakítja, ami alapvetően a magtestecseket körülvevő magalapanyagba lévő molekulák töltésállapotától függ. A kromatin környezetének töltésállapotát az ionok, kivált a magnéziumionok jelentősen befolyásolják.
● A hisztonmagot módosítja az N-végek ubikvitinezése is. Például a H2B 120-as lizinjének társulása ubikvitinnel (ubH2B) megnyújtja az átíródást a FACT (Facilitates Chromatin Transcription) dajkafehérjével együttműködve.
■ A kromatinállapot szabályozásának másik eleme a H1, az összekötő hiszton, amelyik tapad az összekötő DNS két (hisztonmagi) végéhez. Rögzíti a kromatint, gátlására lazul a szerkezet.
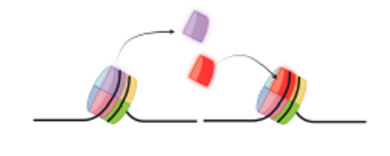 ■ Befolyásolja a
kromatin állapotát maga a DNS is. Például Mg++ hatására megváltozó
DNS a kromatin tömörődését vonja maga után. Az
újabb vizsgálatok kiderítették, hogy a DNS negatív töltése az összekötő
szálakon megmarad, és az összekötő szál töltéskötésekkel kapcsolódhat az
összekötő H1-hisztonhoz vagy más pozitív töltésű fehérjékhez, illetőleg pozitív
ionokhoz. A kromatinállapot szabályozói cserélhetik is a hisztonmag valamelyik
hisztonját. Az ábra két magtestecs egy-egy hisztonjának kicserélődését mutatja.
(Baghy Kornélia észítette.)
■ Befolyásolja a
kromatin állapotát maga a DNS is. Például Mg++ hatására megváltozó
DNS a kromatin tömörődését vonja maga után. Az
újabb vizsgálatok kiderítették, hogy a DNS negatív töltése az összekötő
szálakon megmarad, és az összekötő szál töltéskötésekkel kapcsolódhat az
összekötő H1-hisztonhoz vagy más pozitív töltésű fehérjékhez, illetőleg pozitív
ionokhoz. A kromatinállapot szabályozói cserélhetik is a hisztonmag valamelyik
hisztonját. Az ábra két magtestecs egy-egy hisztonjának kicserélődését mutatja.
(Baghy Kornélia észítette.)
kromatinmódosító össztesek chromatin remodeling complexes a kromatin állapotát úgy megváltoztató többfehérjés (többegységes) össztesek, ami lehetővé teszi, hogy az átírás- és más fehérjék hozzáférjenek a DNS-hez, a sejtfolyamatok (átíródás, DNS-kettőződés, DNS-javítás) bekövetkezzenek. Ezt döntően a magtestecskék mozgatásával érik el. Mindegyiknek van ATPáz gomolya, amely SNF2 ( Sucrose Non- Fermentable 2) gomoly, ezért az SNF2 fehérjék nagycsaládjához tartoznak. Szokásosan egy vagy több kiegészítő gomoly társul hozzá, lehetővé téve egyéb tevékenységeket. Az össztesek mindegyik helikáz, a bázispárok közötti hidrogénkötéseket bontja. ATP felhasználásával. Az SNF2 gomolyuk hasonlósága/eltérése szerint négy családjuk ismert: az →SWI/SNF, az →ISWI, a →CHD és az →INO80.
kromatinmódosulás* (chromatin remodeling) a kromatin olyan átalakulása, ami lehetővé teszi, hogy a DNS-folyamatokat (DNS-átíródás, DNS-kettőződés, DNS-javítás) irányító fehérjék kapcsolódjanak a DNS-hez. A kromatinba zárt DNS tétlen, csak a szabad DNS-szál íródhat át, kettőződhet; például a polimeráz a hisztonhoz kötött DNS-en nem képes végighaladni.
Az átírásfehérjék a magtestecsben lévő DNS-hez nem tudnak kötődni; néhány kivétel azonban van. Az utóbbiakat a nemzetközi irodalom pioneer factorsnek nevezi. Ezek is csak a DNS sajátosan fogékony helyeihez (specific hypersensitive sites, HS sites) kapcsolódva érhetik el a DNS-t.
A kromatin lazítását, a fehérjéknek a hozzáférését kromatinmódosító össztesek végzik a magtestecsek mozgatásával. Például:
▪ A magtestecsek közötti távolság növelése. A DNS rövid szakaszon letekeredik a hisztonmagról, hosszabb lesz az összekötő DNS, így az eleddig a hisztonmaggal zárt DNS egy része is hozzáférhetővé válik.

▪ Egy-egy hisztonmag eltávolítása; a hisztonmagra tekeredett DNS egésze válik szabaddá.

kromatin szerepe az osztódásban A DNS kettőződésével egyidejűleg nagysokaság hiszton képződik a ~20 millió magtestecs felépítésére, amelyekre az átmásolódott DNS rögvest tekeredik. A magtestecsek nagyrészt a régiek újrafelhasználásával jönnek létre. Az osztódási villa széttöri a kromatin, a magtesteceket is. Az újrafelhasználáskor a hisztonpárok (H2A–H2B) megőrződnek.
Átíródási és átíródás után ellenőrző folyamatok biztosítják a keletkező hisztonok megfelelő mennyiségét, ne termelődjék több. Ha nem elegendő, megáll a sejtkör, a többlet pedig zavarja a további lépéseket.
Lényeges a hiszton mag N-végi szabad végződéseinek (hiszton farkok) megfelelő volta: módosulásuknak (metilezés, acetilezés vagy a szerin/treonin foszforilezése) súlyos következményekkel járhat.
kromatinszerkezet A kromatin alapeleme a magtestecs* (nucleosome), amely a hisztonmagból és a köré tekeredet DNS-ből áll. Kapcsolódik hozzá az összekötő hiszton (linker histone); ez nem része csak rögzítője a magtestecsnek.
 • Hisztonmag.
A négyféle alaphisztonból (H2A, H2B, H3 és a H4) tevődik össze. Mindegyikéből
kettő van benne (hisztonnyolcas, histone octamer); először a két H3 és két H4
kötődik egymáshoz, létrehozva a H3–H4 négyest, ehhez tapad két H2A–H2B kettős.
Ezeknek a hisztonoknak a C-végi kétharmada kézfogásszerűen kötődik egymáshoz,
létrehozva a hisztonmagot, amelyből a N-végi gomolyok kitüremkednek.
• Hisztonmag.
A négyféle alaphisztonból (H2A, H2B, H3 és a H4) tevődik össze. Mindegyikéből
kettő van benne (hisztonnyolcas, histone octamer); először a két H3 és két H4
kötődik egymáshoz, létrehozva a H3–H4 négyest, ehhez tapad két H2A–H2B kettős.
Ezeknek a hisztonoknak a C-végi kétharmada kézfogásszerűen kötődik egymáshoz,
létrehozva a hisztonmagot, amelyből a N-végi gomolyok kitüremkednek.
A hisztonoknak a hisztonmagon túlérő N-végeit hisztonfaroknak is nevezik. A H2A-nak a C-vége is túlér, így összesen 10 végződés ér túl a hisztonmagon. A végződések erősen pozitívak a bennük lévő lizin és arginin többség miatt. A kiálló véggomolyok sokféleképpen kapcsolódhatnak fehérjékkel, módosulhatnak (pl. foszforilezéssel, acetilezéssel stb.), további kölcsönhatásba léphetnek a szomszédos magtestecskével is; így befolyásolják a DNS működését, a kromatin szerveződését stb.
Az alaphisztonok helyett sokszor hasonmás hisztonok (H3.3, H2AX, H2AZ, H2ABbd, macroH2A, H2AL1, H2AL2, CENP-A, TH2A, TH2B, H3T, H2BFWT, H3V, H4V, H2BV és H2Abd) képezik a hisztonmagot, ami a kromatin tulajdonságára, a DNS működésére is kihatással van.
▪ DNS. A hisztonmag felszínén a DNS 1,7-szer (~147 bázispár), bal csavarmenet szerint tekeredik körbe, tehát van egy belépő és kilépő szála, folytatódva a szomszédos magtestecskéig (összekötő DNS-szál, linker DNA). A DNS szorosan kapcsolódik a hisztonmaghoz töltéskötésekkel, amelyek a pozitív töltésű aminosavak és a negatív töltésű DNS-váz foszfátcsoportjai között alakulnak ki. A nemzetközi irodalomban a hisztonmag körüli DNS helyzetét dyad positionnak nevezik (hisztonkötött DNS), szemben a magtestecskék közötti összekötő DNS-sel. Az összekötő DNS ~20–80 bázispárnyi (~6,6–27 nm).
• Összekötő hiszton (linker histone). Az emberi sejtekben a H1, ritkábban a H5 hiszton egyetlen molekulája. Szorosan tapad a magtestecshez, rögzíti és tömöríti annak szerkezetét. Az összekötő hiszton a mellette lévő magtestecs összekötő hisztonáig terjed, azokkal egyesülve tengelyt képez, ezen helyezkednek el a magtestecsek az összekötő DNS-sel. Az összekötő hiszton kötődik a hisztonmagból kilépő, illetve belépő DNS-hez, és ezáltal a kromatin tömörödöttségének egyik szabályozója.
kromatinszerveződés hagyományos szemlélet szerint a G0-sejtmagban a DNS és a kromatin szerveződésének három alapeleme fordul elő: 11 nm-es szál, 30 nm-es szál és a 300 nm-es szál.
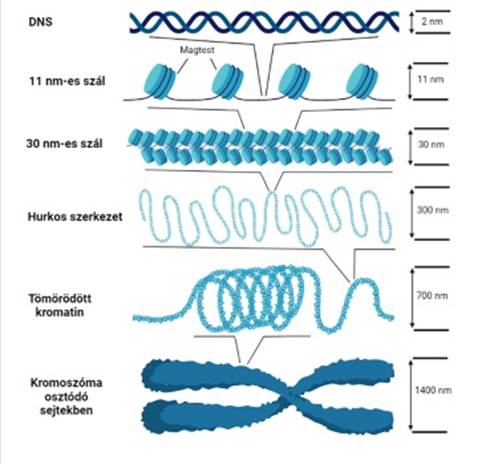 ▪ 11 nm-es
szál (11 nm fiber). A
magtestecsre feltekeredett DNS hosszú vékony szálat képez: ismétlődő
magtestecsek sorozatából és az azokat összekötő DNS-ből (linker DNA) áll. Gyöngyfüzérnek
(„beads-on-a-string”) is nevezik, mert hasonlít rá, de a fonál nem a gyöngyök
közepét átfúrva, hanem azokra feltekeredve halad. A 11 nm-es szál hossza
6–7-szer rövidebb, mint az eredeti DNS-é. A DNS és az összekötő hiszton (H1)
kapcsolata nem tisztázott; a legutóbbi elképzelés szerint a H1 hosszú C-végi
része kötődik a belépő és a kilépő összekötő DNS-sel, ~140 bázispárnyira a
magtestecstől; ez hozza létre a kromatin további szerveződését. A kapcsolódás
rugalmas, foszforilezésre érzékeny: könnyen felszakad, lehetővé téve a kromatin
megnyúlását, a DNS megközelítését.
▪ 11 nm-es
szál (11 nm fiber). A
magtestecsre feltekeredett DNS hosszú vékony szálat képez: ismétlődő
magtestecsek sorozatából és az azokat összekötő DNS-ből (linker DNA) áll. Gyöngyfüzérnek
(„beads-on-a-string”) is nevezik, mert hasonlít rá, de a fonál nem a gyöngyök
közepét átfúrva, hanem azokra feltekeredve halad. A 11 nm-es szál hossza
6–7-szer rövidebb, mint az eredeti DNS-é. A DNS és az összekötő hiszton (H1)
kapcsolata nem tisztázott; a legutóbbi elképzelés szerint a H1 hosszú C-végi
része kötődik a belépő és a kilépő összekötő DNS-sel, ~140 bázispárnyira a
magtestecstől; ez hozza létre a kromatin további szerveződését. A kapcsolódás
rugalmas, foszforilezésre érzékeny: könnyen felszakad, lehetővé téve a kromatin
megnyúlását, a DNS megközelítését.
▪ 30 nm-es szál (30 nm fiber, solenoid) sajátos ún. solenoid szerkezetűnek mondjuk, noha a solenoid szerkezetet sokféleképpen határozták meg, pl. az összekötő hisztonok egymással kötődve tengelyt képeznek, a magtestecsek ezen sorakoznak. Több ilyen képződmény tengelye tömörül úgy, hogy a magtestecsek többfelé helyezkednek el. De olvashatunk olyanról is, hogy a 10 nm-es szál láthatatlan tengely körül tekercselődik, egy-egy menetben ~6 magtestecs van.
▪ 300 nm-es kromatinszál ~100 kilobázisú hurkokat (hurokegységek*) alakító tömörödő kromatinszál. A hurkok részben a maghártyához, részben a magállományhoz rögzülnek. A további képződmények ennek tömörödéséből keletkeznek. (ábra: Baghy Kornélia)
Az új vizsgáló módszerekkel pontosabban tanulmányozható a kromatin szerveződése. A megfigyelések szemléletváltáshoz vezettek: megállapították többek közt, hogy a 30 nm-es szál nem alakul ki az élő sejtben; a kromatin változékony és szabálytalan formákat hoz létre a magtestecsek – főleg töltéshatásokra bekövetkező – mozgása következtében.
Kiderítették, hogy a DNS negatív töltése az összekötő szálakon megmarad, és az összekötő szál töltéskötésekkel kapcsolódhat az összekötő H1-hisztonhoz vagy más pozitív töltésű fehérjékhez, illetőleg pozitív ionokhoz. A maghisztonok pozitív töltései szintén lehetővé teszik a sokrétű kapcsolódásokat a DNS-sel, sőt távoli molekulákkal és a szomszédos magtestecsekkel is. A magtestecsek hisztonvégződéseinek összekapaszkodása a kromatintömörülés egyik alapfolyamata. Bizonyos fehérjék, pl. a kohezin, kifejezetten a magtestecsek társulását, vagyis a kromatin tömörülését okozzák. A kromatin szerveződését tehát a magtestecsek kapcsolódásai, mozgásai alakítják, amelyek alapvetően a magtestecseket körülvevő folyékony magállományban lévő molekulák töltésállapotától függnek.
Végeredményben a kromatin nagyon változékony formájú: a 10 nm-es kromatinszál igen sokféle alakzatba rendeződhet; hasonlóan a folyadékhoz – ezért nevezi a nemzetközi irodalom liquid-like chromatinnak. A kromatin környezetének töltésállapotát az ionok, kivált a magnéziumionok jelentősen befolyásolják.
kromatintömörödés a magtestecsek sürűsödése hozza létre. A tömörített kromatinban a magtestecsek szorosan vannak egymás mellett, rövid az összekötő DNS, és a magtestecssorok sokszorosan hajtódva helyezkednek el, egyre vastagabb szálakat képezve.
A magtestecsek sürösödésében sokminden közrejátszik:
▪ A hisztonmag szabad végeinek acatilezése vagy metilezése, hatócsoportok (foszforilezés, ADP-ribolizáció) kapcsolódása. A metilezés sűrűsíti a magtestecseket, az acetilezés fordítva. A tömött kromatinban kevés az acetilezett hiszton, viszont sok a 9-es lizinen két-háromszorosan metilezett H3 (H3K9me2/3), amelyekhez rendre a tömörítő kromatinfehérjék, például a HP1 (heterochromatin protein 1) kapcsolódik, és toboroz más fehérjéket, hidat képezve a magtestecsek között, tömöríti őket.
Ez a kromatin a késői S-szakaszban másolódik, a számcsökkentő sejtosztódás során nem rendeződik át. Sajátos tulajdonsága, hogy átterjed a szomszédos részekre is, ha a körülmények úgy hozzák. Például, ha a laza kromatin valamely tevékeny génje áthelyeződik a tömörödött kromatinba vagy ahhoz közeli részbe, tétlenné válhat, nem íródik át. Ezt nevezik áttevődéses helyzethatásnak* (position effect variegation, PEV).
A hatócsoportot a hisztonagot módosító enzimek (hisztonmódosító) kapcsolják. Például: a hisztonok acetilezését a hiszton-acetiltranszferáz (histine acetyltransferase, HAT, más néven histone acetylase) végzi: a hiszton N-végi lizinjére acetilcsoportot visz át az acetil-koenzim-A-ról; ε-N-acetil lizin keletkezik.
Az acatilezett hisztonok dezacetilezése ellenkező hatású; tömörödik a kromatin. Az acetilcsoport eltávolításával fokozódik a hiszton pozitív töltése, ezért szorosabban köti a DNS-t, tömöríti a kromatint. A dezacetilezést rendszerint metilezés követi. Például: a H3 acetilcsoportjának eltávolítására a H3 9-es lizinje metileződik; ez némítja a DNS-t, serkenti a kromatin tömörödését. A dezacetilezést a hisztonok ε-N-acetil lizinjének acetilcsoportját eltávolító enzim, a hiszton-dezacetiláz (histone deacetylase, HDAC) sarkallja.
▪ A H1, az összekötő hiszton, amelyik tapad az összekötő DNS két (hisztonmagi) végéhez. Rögzíti a kromatint, gátlására lazul a szerkezet.
▪ Befolyásolja a kromatin állapotát a DNS is. Például a DNS negatív töltése az összekötő szálakon megmarad, és az összekötő szál töltéskötésekkel kapcsolódhat az összekötő H1-hisztonhoz vagy más pozitív töltésű fehérjékhez, illetőleg pozitív ionokhoz, avagy a Mg++ hatására megváltozó DNS a kromatin tömörödését vonja maga után.
▪ Meghatározó a kromatin változása, a maghiszton egyik hisztonjának cserélődése.
▪ Hatással vannak a magtestecsekre a hisztonmaghoz kötődött fehérjék is. Például a bromogomolyt tartalmazó fehérjék elősegítik a kromatin-ATPázok kötődését a hisztonokhoz. Ezek az enzimek elcsúsztatják a magtestecseket, miközben a DNS egy része szabaddá válik, például az indító, és kötődhet hozzá az átírásfehérje.
▪ Módosulnak a magtestecsek ubikvitineződéssel is: lebontódnak, számuk csökken.
Lévén tömörített, a tömörödött kromatin kisebb helyen elfér, ezért azok a sejtmagok, amelyekben a kromatin zöme tömörödött, rendszerint kicsik, zsugorodottak (pycnotic nuclei). A kevésbé zsugorodott magokban a tömörödött kromatin szemcsék formájában látható. Ezek a kromoszómák kromatinná visszaalakulásakor keletkeznek („kromoszóma-maradványok”), vagyis a sejtosztódás származékai.
Összegezve: a laza és a tömörödött kromatinszerkezet alapvetően csak a magtestecsek számában tér el; az utóbbiban sokkal sűrűbben vannak jelen. Mindkettő változékony szerkezet. Vegyi tulajdonságaik eltérnek: a tömörödött kromatin erősebben festődik, több elektront tömörít (elektronmikrográfia), ellenállóbb a DNS-t oldó molekuláknak.
laza kromatin euchromatin a tevékeny kromatin. (→kromatinállapot)
tömör kromatin constitutive heterochromatin (alkati tömörödő kromatin) olyan nem átíródó, ismétletekből, ugrálatokból álló DNS-szakaszok kromatintja, amelyek minden sejtben jelen vannak, kizárólag tömör formában. (→kromatinállapot)
tömörödő kromatin olyan tömörödött kromatin, amelyikben nincs megnyilvánuló átíródás, de külső hatásokra átmenetileg átalakulhat laza kromatinná, és átíródhat. (→kromatinállapot)
tömörödött kromatin a kromatin tétlen formája abban az értelemben, hogy nincs benne génátírás. (→kromatinállapot)
Találatok alcímszavakban (1 szócikk):
hetero- (előtagként) különböző, más, vegyes ■ heterogenous sokrétű, sokféle (különböző részekből összetevődő), egyveleg ■ heterogeneity vegyesség ■ heterokromatin tömörödött kromatin ■ heterolog másféle, különböző.
Részleges egyezések (2 szócikk):
constitutive heterochromatin (konstitutív heterokromatin) tömör kromatin (→kromatinállapot)
DNS-kettőződés és a kromatin átalakulása A DNS kettőződésének a kettőződési villákban szigorúan összhangban kell lennie a buborékban végbemenő kromatin átalakulásával. A DNS-szálak másolódásához a DNS-láncnak el kell válnia a magtestecs hisztonmagjától, ami a magtestecs felbomlásával következik be. A magtestecsek, illetve hisztonok kikerülnek a kettőződési villából, hogy ne akadályozzák a kettőződést. A kettőződést követően, tehát a villa záródásakor azonnal helyre kell állítódnia a kromatinnak. A szabaddá vált hisztonok beépülnek az új megtestecsekbe. Ugyanakkor új hisztonok sokasága is képződik; ezek szintén az új magtestecsek alkotói. Az új hisztonok szintén az S-szakaszban keletkeznek. Az új DNS-láncokhoz azonnal kapcsolódnak a hisztonok, létrehozva az új magtestecseket. Az újonnan képződött hisztonok a késlekedőszálat, az eredeti hisztonok a vezetőszálat tartalmazó DNS-lánchoz kötődnek.
Mint már tárgyaltuk, a magtestecseknek lényeges szerepe van a DNS-kettőződés folyamatának elindításában. A H4 hiszton 20-as lizinjének metilezése, valamint az 5-ös és 12-es lizinjének acetilezése lényeges az előkettőződési össztes kialakításában. Az újonnan képződött H3 és H4 is acetilezett állapotban van az új magtestecs kialakítása előtt.