kromatinállapot A kromatin kétféle állapotban, lazult (laza kromatin) és tömörödött formában van jelen a nyugalmi sejtmagban. Az utóbbinak is két formáját, az állandóan tömörödött (constitutive heterochromatin) és az időszakosan tömörödött (facultative heterochromatin) kromatint különböztetjük meg. A kettő között lényegi különbség van: az állandóan tömörödöttben nincsenek, az időszakosan tömörödöttekben vannak gének, csak némítva.
A lazult forma nemzetközi elnevezése euchromatin, a tömörödötté heterochromatin. Ezt a hazai nevezettár is átvette magyarosan írva: eukromatin és heterokromatin formában. A heterochromatin elnevezés megtévesztő: arra utal, hogy valami más molekula is van benne, noha nincs. Az eu- előtag jelentése ’jól, könnyen’; ellentéte a dys- előtag. Ennek tükrében az euchromatin a jó kromatin. Akkor a heterochromatin a rossz kromatin, ami nem igaz, hiszen igen jelentős szerepe van a sejt működésében. A fentiek miatt a laza kromatin és a tömörödött kromatin elnevezés kifejezőbb, megfelelőbb. Mivel az állandóan és időszakosan tömörödött kromatin között lényegi különbség van: célszerű külön névvel illetni őket. Az állandóan tömörített kromatinnak tömör kromatin*, az időszakosan tömörödöttnek tömörödő kromatin* lehet a neve.
■ A laza kromatin a tevékeny kromatin. Génszerkezeteket tartalmaz, és lehetővé teszi azok átíródását: a laza szerkezete következtében az átírásfehérjék, enzimek, RNS-ek hozzáférnek a DNS-hez. Jellegzetes a hisztonvégek (histone tails) acetilezett állapota. A magtestecsek egyvonalban sorakoznak, gyöngyfüzér alakzatban vannak; a köztük lévő távolság viszonylag nagy, miként az összekötő DNS is, több a hozzáférhető hely.
A laza kromatin a kromatinállomány ~90%-át teszi ki, főleg a sejtmag belsejében helyezkedik el. A tevékeny sejtmagokban sok a laza kromatin, és mivel helyigényes, hiszen kevésbé tömörített, a sejtmagok nagyobbak. A laza kromatin a sejtosztódás korai S-szakaszában másolódik.
■ Tömörödött kromatin a kromatin tétlen formája abban az értelemben, hogy nincs benne génátírás. A kétféle tömörödött kromatin, a tömör és a tömörödő kromatin szerkezete és jelentősége is eltérő.
▪ Tömör kromatin olyan kromoszómarész, amelyik minden sejtben jelen van, kizárólag tömörödött formában; ismétletekből és ugrálatokból tevődik össze, nem íródik át. A kromoszómák alapvető rögzítő szerkezeti eleme, a befűződések és a végrészek körüli DNS-t alkotja. Jellemzi a sokalakúság, és hogy C-sávot tartalmaz. Bizonyos sejtekben sajátos helyzetű. A nyugalmi sejtmagban összenyomott, sötéten festődő szemcsék formájában látható.
▪ Tömörödő kromatin olyan tömörödött kromatin, amelyikben nincs megnyilvánuló átíródás, de külső hatásokra átmenetileg átalakulhat laza kromatinná, és átíródhat. Jellemzően hosszú elszórt ugrálatokat tartalmaz, és vannak benne gének is, de nincs C-sáv. Meglehetősen egyforma, de változékony szerkezetű, nem sokalakú. A néma X-kromoszómát ez alkotja. Feltételezhető, hogy a sejtfejlődés folyamán azok a gének, amelyek az elődsejtekben tevékenyek, de az érett sejt működésében már szükségtelenek, tömörödő kromatin képződésével némulnak el.
A tömörödött kromatinban kevés az acetilezett hiszton, viszont sok a 9-es lizinen két-háromszorosan metilezett H3 (H3K9me2/3), amelyekhez rendre a tömörödött kromatinfehérjék, például a HP1 (heterochromatin protein 1) kapcsolódik, amely más fehérjéket toboroz, és hidat képezve a magtestecsek között, tömöríti őket. A tömörített kromatinban a magtestecsek szorosan vannak egymás mellett, rövid az összekötő DNS, és a magtestecssorok sokszorosan hajtódva helyezkednek el, egyre vastagabb szálakat képezve.
Ez a kromatin a késői S-szakaszban másolódik, a számcsökkentő sejtosztódásban nem rendeződik át. Sajátos tulajdonsága, hogy átterjed a szomszédos részekre is, ha a körülmények úgy hozzák. Például, ha a laza kromatin valamely tevékeny génje áthelyeződik a tömörödött kromatinba vagy közeli részbe, tétlenné válhat, nem íródik át. Ezt nevezik áttevődéses helyzethatásnak* (position effect variegation, PEV).
Lévén tömörített, a tömörödött kromatin kisebb helyen elfér, ezért azok a sejtmagok, amelyekben a kromatin zöme tömörödött, rendszerint kicsik, zsugorodottak (picnotic nuclei). A kevésbé zsugorodott magokban a tömörödött kromatin szemcsék formájában látható. A kromoszómák kromatinná visszaalakulásakor keletkezik („kromoszóma-maradványok”), vagyis a sejtosztódás származékai.
A laza és a tömörödött kromatin szerkezet alapvetően csak a magtestecsek számában tér el; az utóbbiban sokkal sűrűbben vannak. Mindkettő változékony szerkezet. Vegyi tulajdonságaik azonban eltérnek: a tömörödött kromatin erősebben festődik, több elektront tömörít (elektronmikrográfia), ellenállóbb a DNS-t oldó molekuláknak.
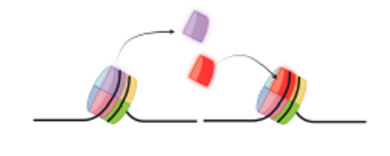 ■ Befolyásolja a
kromatin állapotát maga a DNS is. Például Mg++ hatására megváltozó
DNS a kromatin tömörődését vonja maga után. Az
újabb vizsgálatok kiderítették, hogy a DNS negatív töltése az összekötő
szálakon megmarad, és az összekötő szál töltéskötésekkel kapcsolódhat az
összekötő H1-hisztonhoz vagy más pozitív töltésű fehérjékhez, illetőleg pozitív
ionokhoz. A kromatinállapot szabályozói cserélhetik is a hisztonmag valamelyik
hisztonját. Az ábra két magtestecs egy-egy hisztonjának kicserélődését mutatja.
(Baghy Kornélia észítette.)
■ Befolyásolja a
kromatin állapotát maga a DNS is. Például Mg++ hatására megváltozó
DNS a kromatin tömörődését vonja maga után. Az
újabb vizsgálatok kiderítették, hogy a DNS negatív töltése az összekötő
szálakon megmarad, és az összekötő szál töltéskötésekkel kapcsolódhat az
összekötő H1-hisztonhoz vagy más pozitív töltésű fehérjékhez, illetőleg pozitív
ionokhoz. A kromatinállapot szabályozói cserélhetik is a hisztonmag valamelyik
hisztonját. Az ábra két magtestecs egy-egy hisztonjának kicserélődését mutatja.
(Baghy Kornélia észítette.)