aligkarú kromoszómák acrocentric chromosomes azok a kromoszómák, amelyeknek a befűződése csaknem a rövid kar végén van, kicsik. Ilyenek: a 13, 14, 15, 21 és 22, valamint az Y-kromoszóma.
Találatok címszavakban (3 szócikk):
kromoszómák alapvető festési módszerei Négyféle van: Giemsa-festés (G-sávozás), Q-sávozás (Q-banding), R-sávozás (R-banding) és a C-sávozás (C-banding).
Forrás: https://www.nature.com/scitable/topicpage/karyotyping-for-chromosomal-abnormalities-298/
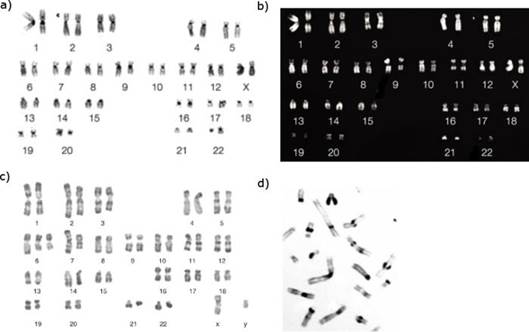
a) Giemsa-festés (G-sávozás). A G-sávozáskor az osztódásos (metaphase) kromoszómákat fehérjebontó enzimmel, például tripszinnel, kezeljük, majd Giemsa-festéket adunk hozzá. A festék a fehérjebontást követően a DNS bázispárjai közé ékelődik be. Az AT-ben (adenin-timinben) gazdag részek sötét sávokként lesznek láthatóak, míg a GC-ben (guanin-citozinben) gazdag területek világos csíkok formájában jelennek meg. Ennek oka, hogy a tripszin a kromoszómák átírásban tevékeny, kevésbé tömörödött részein – ezek a GC-ben bő részek – a fehérjékhez könnyebben hozzáfér, többet emészt meg, ezért kevesebb Giemsa-festék kötődik a DNS kettősszálhoz, így világosabb sáv keletkezik.
A G-sávozással kitűnően tájékozódhatunk a kromoszómák belső szerkezetéről; a kromoszómahelyek egyik meghatározási módja, például a gének helyét a kromoszómák számának, karjának és G-sávjainak megadásával írjuk le.
b) Q-sávozás (Q-banding) szintén az AT- és a GC-bőség szerint fest. Itt is a közbensőszakasz (metaphase) kromoszómákat festjük, de fluoreszcens festéket (például quinacrine) alkalmazunk, és fluoreszcens mikroszkóppal értékelünk. A kapott sávozás fordított: itt az AT-ben gazdag kromoszóma részek lesznek világosak, feltűnőek, míg GC-ben gazdag területeken fakó sávok keletkeznek. Ennek oka az, hogy csak az AT-quinacrine együtt ad fluoreszcens jelet, a GC-quinacrin kötődés kioltja a fluoreszcens festék jelét. A fluoreszcens jel egyenesen arányos az AT bázispárok mennyiségével. A módszer alkalmas az Y kromoszóma és számos sokalakúság (például ismétletek) kimutatására.
c) R-sávozás (R-banding) a G-sávozás kiegészítője. A kromoszómák végén előforduló eltéréseket (például törlődések, áthelyeződések), illetve a génben gazdag területeket lehet ezzel a festéssel különösen jól megjeleníteni, ugyanis ezek G-festéssel gyakran gyengén festődnek.
Először a DNS két szálát szétválasztják forró savas sóoldattal, majd Giemsa-festék használatával sávokat kapunk. A G-sávozással ellentétben, az AT-ben gazdag kromoszóma sávok kapnak világos színt, míg a GC-ben gazdag részek sötét színt vesznek fel.
d) C-sávozás (C-banding): a kromoszómák tömörített kromatint tartalmazó területeinek a részleteit tanulmányozhatjuk vele. Szintén Giemsa -festéket alkalmazunk, melyet savval és bázissal való kezelés előz meg. A savas kezelés során a kettősszálú DNS szétválik, a bázikus kezelés hatására az egyszálú DNS-ek ismét bázispárosodnak. A módszer ezeknek a folyamatoknak a gyorsaságán alapszik.
Leginkább a tömör (constitutive heterochromatin) kromatint, tehát ismétlődő, nem kódoló elemeket (ugrálatok, különböző ismétletek) tartalmazó DNS-szakaszok elemzésére alkalmazzuk. Ilyenek vannak a befűződésekben, végrészeken, azokkal szomszédos területeken és az 1., 9., 16., 19. és Y kromoszómában.
Az ismétlődő szakaszokat tartalmazó DNS-részek gyorsabban festődnek Giemsa-festékkel.
kromoszómák kiegészítő festési módszerei Kétfélét alkalmazunk.
▪ FISH (fluorescent in situ hybridization). A FISH vizsgálatot alakított kromoszómákon, sejteken, lenyomati vagy egyéb szöveti metszeteken is el lehet végezni, de visszérből, köldökvérből, amnionból vett mintákban (amnionvíz, chorionminta), nyiroksejteken és paraffinba ágyazott mintákból is.
A módszer rövid, fluoreszcensen jelölt DNS/RNS mintákat alkalmaz, amelyek bázispárosodnak a kromoszómán.
A minták előkészítése azok fajtájától függ. Pepszinnel emésztik a szövet fehérjéit. Ezután a kromoszómák és a minták szálait szétválasztják, és a mintát a fluoreszcens mintával hőn tárolják 12-24 órán keresztül. Ez idő alatt a minta a kiegészítő DNS szakasszal kereszteződik (amennyiben van olyan a kromoszómán). A sejtmagokat DAPI festékkel teszik láthatóvá, végül fluoreszcens mikroszkóp segítségével vizsgálják a folyamat eredményét.
A FISH jelentősége abban van, hogy nagyon gyorsan mutatja ki a kórismét megerősítő elváltozást, ami, például válságos állapotban lévő csecsemőnél nagyon fontos.
Használható:
- számbeli kromoszóma-eltérés kimutatására (például háromféle kromoszóma kimutatására fajlagos mintát alkalmazunk. Ha látható mind a három, igazolt a kromoszómahármas),
- mikrotörések (például valamely és a mellette lévő G-sávra fajlagos a DNS-minta; ha egyik nem festődik, a sáv hiányára utal),
- bonyolult áthelyeződések (olyanokra is, amelyek G-sávokkal nem azonosíthatók),
- de gének túlkifejeződésére is.
▪ m-FISH / multicolor FISH: lépései nagyjából megegyeznek a FISH-ével, csak egyszerre akár 24 különböző fluoreszcens DNS/RNS próbát alkalmazhatnak.
Részleges egyezések (2 szócikk):
acrocentric chromosomes →aligkarú kromoszómák
kromoszómapár chromosome pair A kromoszómák párokba rendeződnek; egy-egy párt egy anyai és egy apai kromoszóma alkot. Elkülönítünk 22 testi kromoszómapárt (44 testi kromoszóma) és egy nemi kromoszómapárt, amely nőkben XX-, férfiakban XY-kromoszómákból áll. Az utóbbiak lényegileg eltérők, de az XX-pár sem azonos, mert az egyik X némított. Ezzel szemben a testi kromoszómák egyformák: ugyanazon a helyeken, ugyanazokat a tulajdonságokat hordozzák.
A kromoszómapárokat 1-től 22-ig számozzuk. Vizsgálatakor a párokat sorba rakjuk az 1-essel kezdve; a testi kromoszómák után tesszük a két nemi kromoszómát. Az így kirakott kromoszómasornak karyotype a nemzetközi neve; magyarul kromoszómakép.
A kromoszómapárok tagjait az
angol irodalom homologous chromosomes (homologs)-nak nevezi. Magyarul
rokonmás kromoszómák. A hazai irodalom is használja a homológ kromoszómák
szakszót, de szükségtelenül. A kromoszómapár teljesen megfelelő, és
mondható a rokonmás kromoszóma is.