lánc a vegytanban egyféle molekula, egyetlen egységsorból álló nagymolekula, például polipeptid (peptidlánc), aminosavlánc. Nanométer (nm) nagyságú.
Találatok alcímszavakban (1 szócikk):
szerkezeti azonmások* constitutional isomers azonos atomösszetételű és molekulatömegű, de az atomok kapcsolódásában (összekapcsolódási rendjében) eltérő molekulák. Másként: azonos összegképletű, de különböző szerkezeti képletű vegyületek. A szerkezeti azonmások fizikai tulajdonságai, pl. forráspontjuk, különbözik. (→azonmás)
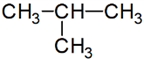
lánc azonmások* chain isomers az atomok, szokásosan a szénatom elágazásában és/vagy kötésükben eltérő azonmások. Pl.
sorrendi eltérés:
n-bután:
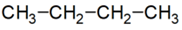
izobután:
kötési eltérés:
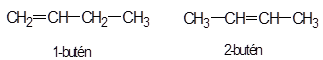
A legegyszerűbb szénhidrogéneknek, mint metán (CH4), etán (CH3–CH3) és propán (CH3–CH2–CH3), nincs azonmása, mert a szén és hidrogének kapcsolódása csak egyféleképpen lehetséges.
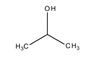
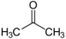 hatócsoportú
azonmások* functional (group) isomers
különböző hatócsoportot kapcsoló azonmások. Pl. az acetonban karbonil- (C=O), a
propanolban pedig hidroxilcsoport van.
hatócsoportú
azonmások* functional (group) isomers
különböző hatócsoportot kapcsoló azonmások. Pl. az acetonban karbonil- (C=O), a
propanolban pedig hidroxilcsoport van.
kötésváltó azonmások* tautomers csak a protonok és elektronok elhelyezkedésében más azonmások. A kötésátalakulás voltaképpen protonáthelyeződés* (prototropy): egy proton a molekula egyik helyéről a másikra tevődik át, rendszerint az elektronok eloszlási változásával, a mozgékony hidrogén áttevődésével és kötésváltozással jár. Ezek egyféle kötésben eltérő vegyületek, amelyek könnyen átalakulnak egymásba. Együtt vannak jelen, váltakozó egyensúlyi állapotban, és a vegyi folyamatokban szintén együtt vesznek részt. A kötésváltásnak* (tautomerism) számos formája ismert. Néhány gyakoribb:
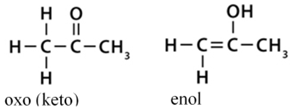 • oxo(keto)–enol azonmásság*. Az α helyzetű szénatom hidrogénjének
áthelyeződése. Az oxovegyületekben egy oxigénatom kettős kötéssel kapcsolódik
egy szénatomhoz. Az enolformában OH-csoport kötődik telítetlen szénvegyülethez.
Az oxovegyületek előfordulhatnak
enolformában is (oxo–enol kötésváltás). Az oxoforma erősebb kötésű, és túlsúlyban van a kettő egyensúlyi
állapotában. (→hatáscsoport, oxovegyületek)
• oxo(keto)–enol azonmásság*. Az α helyzetű szénatom hidrogénjének
áthelyeződése. Az oxovegyületekben egy oxigénatom kettős kötéssel kapcsolódik
egy szénatomhoz. Az enolformában OH-csoport kötődik telítetlen szénvegyülethez.
Az oxovegyületek előfordulhatnak
enolformában is (oxo–enol kötésváltás). Az oxoforma erősebb kötésű, és túlsúlyban van a kettő egyensúlyi
állapotában. (→hatáscsoport, oxovegyületek)
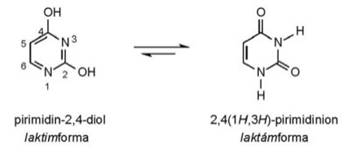 • laktim–laktám azonmások. Az α helyzetű nitrogénatom
hidrogénjének áthelyeződése: –N=C– / –N–C–;
a laktimban a szén kettős kötéssel, a laktámban egyes kötéssel kapcsolódik a
nitrogénhez. Gyűrűs vegyületekben fordul elő, a
gyűrűs savamidokra (laktámokra) jellemző a hidrogénátrendeződés a
gyűrűben lévő amidcsoporton belül. Az uracil
(pirimidin-2,4-diol) egyensúlyi folyamata:
• laktim–laktám azonmások. Az α helyzetű nitrogénatom
hidrogénjének áthelyeződése: –N=C– / –N–C–;
a laktimban a szén kettős kötéssel, a laktámban egyes kötéssel kapcsolódik a
nitrogénhez. Gyűrűs vegyületekben fordul elő, a
gyűrűs savamidokra (laktámokra) jellemző a hidrogénátrendeződés a
gyűrűben lévő amidcsoporton belül. Az uracil
(pirimidin-2,4-diol) egyensúlyi folyamata:
• Egyéb kötésváltó azonmások: enamin–imin: H−N−C=C / N=C−C−H, nitrozo–oxim: H−C−N=O / C=N−O−H, keten–inol: H−C=C=O / C≡C−O−H, foszfit–foszfonát: P(OR)2(OH) ⇌ HP(OR)2(=O), aminosav–ammonium-karboxilát (H2N−CH2−COOH / H3N+−CH2−CO−) stb.
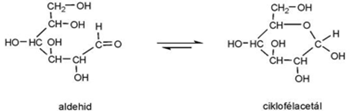 gyűrű–lánc azonmások* ring-chain isomers az azonmások egyike nyíltlánc, a másik gyűrűs. Olyan
aldehid-/ketocsoportot és OH-csoportot tartalmazó vegyületekben, például
szénhidrátokban fordul elő, amelyekben a proton áthelyeződésével a nyílt
vegyületből (láncforma) gyűrűs lesz, és fordítva. Az OH-csoport hidrogénje az
aldehidcsoportra vándorol, kialakítva a gyűrűformát. Az aldehidlánc utolsó
szénatomjához kettős kötéssel kapcsolódó oxigén helyett egyes kötéssel
OH-csoport társul. Az ábrán a glükóz (szőlőcukor) példája látható.
gyűrű–lánc azonmások* ring-chain isomers az azonmások egyike nyíltlánc, a másik gyűrűs. Olyan
aldehid-/ketocsoportot és OH-csoportot tartalmazó vegyületekben, például
szénhidrátokban fordul elő, amelyekben a proton áthelyeződésével a nyílt
vegyületből (láncforma) gyűrűs lesz, és fordítva. Az OH-csoport hidrogénje az
aldehidcsoportra vándorol, kialakítva a gyűrűformát. Az aldehidlánc utolsó
szénatomjához kettős kötéssel kapcsolódó oxigén helyett egyes kötéssel
OH-csoport társul. Az ábrán a glükóz (szőlőcukor) példája látható.
Részleges egyezések (6 szócikk):
chain →lánc ■ chain isomerism lánc azonmásság (→szerkezeti azonmásság)
jelközvetítés* signaling pathway a jelzés továbbadásának sejten belüli folyamata. Az idegsejtek ingerületvezetésétől eltekintve, molekulák végzik; a jelfogótól a végrehajtó molekula keletkezéséig vagy tevősítéséig, a jelzésvégrehajtásig terjed. Ha a jelátadás már önmagában sejtválaszt eredményez, például megnyílik az ioncsatorna, a jelútnak ez a része kimarad. Ha a jelátadás közvetlenül átírásfehérjét tevősít, a jelközvetítés többnyire egymolekulás: az átírásfehérje bejut a sejtmagba, közvetve vagy közvetlenül kötődik a génhez, és elindítja a sejtválaszt – ez jellemző például a szteroidok jelközvetítésére. Legtöbbször azonban fehérjék stb. gerjesztődnek egymás után foszforileződéssel vagy csupán két fehérje társulására létrejövő térszerkezeti változás következtében.
jelközvetítési láncfolyamat Szokásosan a jelátvitellel gerjesztett enzim másik fehérjét tevősít; ezt sejtbeli hírvivőnek*, (intracellular messenger) mondjuk. Avagy valamilyen gyorsan terjedő kismolekula (például cAMP) vagy ion (kalcium) megjelenésével jár; ez utóbbiakat másodlagos hírvivőnek* (secondary messenger) nevezzük. Előfordul, hogy ugyanaz a másodlagos hírvivő többféle jelátadás (más-más jelvivő–jelfogó kapcsolat) hatására is megjelenik a sejtben.
A másodlagos hírvivő vagy a sejtbeli hírvivő további fehérjé(ke)t, enzime(ke)t gerjeszt, ezek megint újabbat és így tovább, vagyis a folyamat ismétlődő lépésekből áll, ezért nevezzük láncfolyamatnak. A folyamatban szereplő molekulák a jelközvetítők* (intracellular mediators). Előfordul, hogy valamely tevősített jelközvetítő a sejt egyik részéből, akár távolabbi pontról is, a másikba, leginkább a sejthártyához vándorol, hogy ott kapcsolódjék másik fehérjével – egy-egy láncszem csupán ezért iktatódik be.
A jelközvetítés a végrehajtó molekula (általában átírásfehérje) gerjesztésével zárul. A folyamat nagyon gyors, és teljesen szabályozott. Egy-egy láncfolyamat tehát a jelfogó bekapcsolásától sejtbeli fehérjék egymást követő tevősítésén át a végrehajtó molekuláig terjed. A láncnak bármely tagja többféle enzimet is bekapcsolhat, így a folyamat több irányban is haladhat, de egyszerre rendszerint csak egy irányban megy végbe.
A jelközvetítés az enzimláncon keresztül tehát meglehetősen bonyolult és szerteágazó lehet. Ennek számos előnye van:
• Egyfajta sejtműködést, például a cukorfelvételt, többszörös jelközvetítéssel, sőt többféle jelvitellel is biztosítja a sejt. Ezek között külön-külön és együttesen is összeköttetés (cross talk) van, egyébként nem valósulhat meg az összehangolt szabályozás. Ez a lényegi különbség a jóval egyszerűbb vagy akár csak egyetlen molekulával végbemenő jelközvetítéssel szemben. Például: ha az inzulin jelútban közvetlen kapcsolat lenne a jelfogó és a glikogén-foszforiláz között, a Ca2+ nem kapcsolódhatna be a szabályozásba, azaz a sokoldalú irányítás nem lenne lehetséges. (→inzulinjelút)
• A közvetítések elágazásai további biztonsági, ellenőrző utakat hoznak létre.
• Lehetővé teszi a fehérjekapcsolatok kétirányú kölcsönhatását, azaz, hogy a folyamat oda-vissza is mehet, elősegítve a rugalmas szabályozást.
• Ugyancsak a soklépcsős láncfolyamattal valósítható meg az, hogy a jel a sejt több pontját is elérje, vagy eljusson az egyik szervecskétől a másikba stb.
• Egy-egy többfelé ágazó jelközvetítés rendszerint sokféle sejtfolyamatot vált ki. Ha ezek külön-külön jelközvetítéssel mennének végbe, jóval több molekula részvételére lenne szükség és jóval körülményesebb lenne – a láncfolyamat tehát „takarékos” is - a törzsfejlődés folyamán alakult ilyenné.
A jelzés a jelközvetítéssel általában felerősödik azáltal, hogy minden lépéssel több és több célmolekula gerjesztődik. Például: egyetlen adenilát-cikláz nagyon sok cAMP-t képez; ezek pedig sok-sok fehérje-kinázt serkentenek, azok még több foszforilázt stb. Vannak azonban olyan jelközvetítések is, amelyekben egy molekula csak egy célmolekulával találkozik, úgymond „dobozba (cassette) zárt”.
jelközvetítők zömében enzimek, de kapcsoló- cserélő- és állványfehérjék, vagy más feladatuk is lehet. Az állvány- és a kapcsolófehérjék bevonásának a célja a jelátadónak és célmolekuláinak az összehozása. A jelközvetítésben legtöbbször G-fehérjék vagy más néven GTP-ázok a kapcsolófehérjék: bekapcsolt állapotukban GTP-t kötnek; ennek segítségével foszforilezik az első jelközvetítő fehérjét. A cserélőfehérjék pedig a G-fehérjék bekapcsolásában vesznek részt, segítik a GDP leválását, lehetővé téve a GTP kötődését – mert a G-fehérjék maguk erre nem képesek. A jelközvetítés enzimei leginkább fehérje-kinázok (foszfátcsoportot szállítók); ezek a tirozin vagy a szerin, illetőleg a treonin oldalláncát foszforilezik ATP felhasználásával. A szerin-treonin-kinázok a treoninra és a szerinre is képesek foszfátot kapcsolni, ami az így módosított enzimet serkentheti vagy gátolhatja is, az adott enzimtípustól függően. Gyakoriak az ATPázok, GTPázok, valamint a foszfatázok (levágnak a célmolekuláról foszfátcsoportot [PO43-], defoszforilezés) és a foszforilázok (foszfátiont kapcsolnak ATP felhasználása nélkül), továbbá másodlagos hírvivőként a cAMP (cyclic adenosine monophosphate), cGMP (cyclic guanozin monophosphate), diacilglicerol (DAG), a zsírsavak, az inozitol-triszfoszfát (IP3), sejthártyához kötött foszfoinozitidek és a Ca-ionok. A nitrogén-monoxid (NO) bizonyos értelemben szintén tekinthető másodlagos hírvivőnek is, hiszen a sejtben is viszi a jelzést.
jelszakasz* a jelközvetítés valamely része.
nyílt láncú vegyület open-chain (acyclic, aliphatic) compound egyenes vagy elágazó láncot alkotó szerves vegyület. (→szerves vegyület)
polimeráz polymerase nukleinsavat (RNS-t vagy DNS-t) képező enzim, nukleotidokat kapcsol össze foszfodiészter-kötéssel. A polimerázok több alegységből álló fehérjék; alegységeik fajtánként némileg eltérnek. Általános értelemben a polimeráz soktagú molekulát (polymer) képező enzim. Sokféle polimeráz ismert; két nagy csoportjuk a mintafelülettől függő (DNS-, RNS-polimeráz) és független polimeráz (pl. Poli-A-polimeráz). A DNS-polimeráz DNS-t másol a DNS-ről, az RNS-polimerázok pedig különböző RNS-t készítenek a DNS-szálról, vagy RNS-t másolnak. (→DNS-polimeráz, RNS-polimeráz)
polimeráz-láncfolyamat (PCR) polymerase chain reaction, PCR laboratóriumi módszer valamely DNS-szakasz sokszorosítására. Veleje: a kétszálú DNS-szakasz szétválasztása, a cél DNS-szakasz kijelölése rövid indítószálakkal (primerek), és DNS-képzés az egyszálú kijelölt DNS-szakasz polimerázzal való másolásával. A folyamat nagyon gyors, és sokszorosan ismétlődik, másolatok sokasága keletkezik.
szerves vegyületek organic compounds szénhidrogének; szén- és hidrogénatomokból épülnek fel, de lehetnek bennük más atomok is (oxigén, nitrogén, kén, foszfor stb.), amelyeket idegen atomoknak nevezünk. Az atomok elektronkötéssel kapcsolódnak egymáshoz. A szerves vegyületek egyedülálló tulajdonsága, hogy más szénatomokkal, vagy a szerves vegyületekben előforduló más atomokkal, változatos szerkezeteket (láncok, elágazások, gyűrűk stb.) hoznak létre. Ezek alkotják az élő szervezetet. A szerves vegyületek száma szinte határtalan, több millió ismert, szemben a néhány százezer szervetlen vegyülettel. A természetben előfordul néhány olyan szénatomos vegyület, amelyet nem tartunk szervesnek; ilyenek a karbidok, karbonátok és a cianidok. (→vegykötések)
A szerves vegyületeket háromféle tulajdonság alapján osztályozzuk: aromás és nem aromás, nyíltláncú és gyűrűs, valamint telített és telítetlen szerves vegyületekre.
aromás vegyületek aromatic compounds síkalkatú, egyes és kettős kötésű, hat szénatomos állékony gyűrűs (benzolgyűrű) vegyületek jellegzetes elektronszerkezettel. A kötések mindegyike részt vesz az elektronok mozgásában: 3 pár π-elektron egyenletesen kering minden atom körül (a Hückel-szabály szerint). Az aromás vegyületek telített vegyületként viselkednek; helyettesítő vegyülékenységűek, tulajdonságaik merőben mások, mint az azonos összetételű nyílt láncúaké. (→Hückel-szabály)
gyűrűs (zárt láncú) vegyületek closed (cyclic, ring) compounds szénatomokat tartalmazó gyűrűt képező vegyület, az
első és az utolsó szénatom kapcsolódik egymáshoz. Lehetnek egygyűrűs
(monocyclic compound), kétgyűrűs (bicyclic compound), háromgyűrűs (tricyclic
compound), néhánygyűrűs (oligocyclic compound), sokgyűrűs (polycyclic compound)
összetételűek. A gyűrűk közös atomja szerint lehetnek: 1. elkülönült többgyűrűs
– nincs közös atom; 2. egy közös atomú; 3. két közös atomú és 4. áthidalt
gyűrűs – több közös atomú – vegyületek. Megkülönböztetünk csak szénatomos
gyűrűs vegyületeket (homocyclic [isocyclic] compounds] – a gyűrű valamennyi
tagja szénatom (homociklusos vegyület) – és vegyes gyűrűs vegyületeket
(heterocyclic compounds) – a gyűrűben a szénatomok mellett más atom is van (heterociklusos
vegyület). Mindegyik lehet aromás és nem aromás.
nem aromás vegyületek (alifás vegyületek) minden olyan szerves
vegyület, amelyik nem aromás tulajdonságú. Lehet nyílt láncú és gyűrűs
(alicyclic compounds), valamint telített és telítetlen.
nyílt láncú vegyületek open-chain (acyclic, alophatic) compounds elektronkötésekkel kapcsolódó szénatomok láncából álló szerves vegyületek. A lánc lehet egyenes (linear) és elágazó (branching). Egyszeres (C–C), kettős (C═C) és hármas (C≡C) kötésű szénatomokat (C) tartalmazhatnak.
telítetlen vegyületek unsaturated compounds legalább egy kettős vagy hármas kötéssel kapcsolódó szénatomokat tartalmazó vegyületek. Vegyülékenyek: a kettes/hármas kötés könnyen cserélődik egyesre. Főleg egyesülnek és sokszorozódnak.
telitett vegyület saturated compound a szénatomok mindegyike egyes kötéssel kapcsolódik, nincs szabad kötőhely. Kevéssé vegyülékeny molekulák; főleg helyettesítési folyamatokban vesznek részt. Lehetnek nyílt- és zártláncúak.
zárt láncú (gyűrűs) vegyületek closed (cyclic, ring) compound szénatomos gyűrűt tartalmazó vegyületek. (→szerves vegyületek)