magjelfogók nucleal receptors a sejtben lévő jelfogók; jelvivőt kötő átírásfehérjék a sejtelkülönülés, az ébrényi fejlődés, a szervkialakulás irányítói. A hatásukat a DNS-ben fejtik ki, ahhoz fűződnek – ezért nevezzük ezeket magjelfogóknak –, jóllehet jó néhánynak van sejtfelszíni hatása is. Nagycsaládot alkotnak, amelynek 48 tagja van; 48 különböző gén kódolja őket.
A jelvivőik két nagy csoportba sorolhatók: a koleszterinalapú és a zsírsav/foszfolipid alapú jelvivőkre.
■ A koleszterinalapú jelvivők a szteroid hormonok; ezek kis molekulák, könnyen átjutnak a sejthártyán, és nagy fogékonysággal kötődnek a jelfogókkal, amelyeket hormonjelfogóknak (endocrine receptors) nevezünk. Közéjük tartoznak a mineralo-/glükokortikoid, ösztrogén-, androgén-, progeszteronjelfogók stb.
A sejtplazmában lévő jelfogók dajkafehérjével (pl. HSP90, ritkán HSP70) kapcsolódva vannak, tevőtlen állapotban. A jelvivővel kötődve elválnak a dajkafehérjétől, azonnal kettőst képeznek, pl. két jelvivős ösztrogénjelfogó társul; ez szükséges a tevősödéshez. Kettősként áthatolnak a maghártyán, és kötődnek a célgénhez, a fajlagos hormonkötő válaszelemhez (HRE; hormone response element), amely rendszerint az indítóban vagy a fokozóban van. (→hormonkötő válaszelemek)
A génnel kapcsolt jelfogó–jelvivő kettőshöz társserkentők (transcriptional coactivators, CoA) csatolódnak; többféle kapcsolódik, rendre nem egyszerre, hanem amikor az egyik elvégezte a feladatát, kötődik a másik; avagy az egyik kapcsolja a másikat, pl. az SRC1–3 elsőként járul a jelfogóhoz, és toborozza a többit. Ezek módosítják a kromatint, lehetővé téve az átírásfehérjék kapcsolódását. A társserkentőkben hiszton-acetil-transzferáz van, amely a hisztonok acetilezésével megnyitja a kromatint.
A szteroid hormonok általi génátírás legtöbbször két lépésben zajlik: először a korai géneket serkentik, amelyekről szabályozó fehérjék képződnek. A szabályozó fehérjék egyik csoportja gátolja a korai gének további átírását, másik csoportjuk serkenti a késői géneket, amelyek a hormonhatást végrehajtó fehérjéket kódolják. Ez a kétlépéses átíródás magyarázza az elhúzódó hormonhatást.
■ A zsírsav/foszfolipid alapú jelvivők a sejtmagban lévő jelfogókkal társulnak, szerkezeti változást hoznak létre, a jelfogók így tevősödnek. A sejtmagban lévő jelfogók (pl. pajzsmirigyhormon-, retinolsavjelfogó), társgátlóval (CoR, corepressor) kapcsolódva kötődnek a fajlagos hormonkötő válaszelemhez, tevőtlenek. A jelvivővel csatlakozva elválnak a társgátlótól, egyesülnek az RXR-rel (retinoid X receptor), azaz vegyes kettőst képeznek; így kötődnek a célgénhez. A társgátlók (pl. NCoR, SMRT) hiszton-dezacetilázt (HDAC) kötnek, amely tömöríti a kromatint.
Sok ilyen jelfogó jelvivője nem tisztázott (árva jelfogók, orphan receptors), vagy csak később vált ismertté (árvátlanodott jelfogók (adopted orphan receptors), ezért nevükben előfordulhat az orphan jelző.
Ebbe a csoportba tartozó jelfogók pl. az SF1 (steroidogenic factor-1), a RORα (RARrelated orphan receptor α), a RORγ (retinoic acid receptor-related orphan receptor γ), a D-vitamin-jelfogó stb. Előfordul, hogy az árva jelfogó kettőst képez valamelyik szokványos magjelfogóval; ilyen pl. az RXR (retinoid X receptor).
Néhány sejtmagi jelfogó a sejtplazmában keletkezett jelvivővel társul, lehetővé téve a sejt önszabályozását. Pl. az LXR az oxiszterollal kapcsolódik.
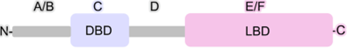 A magjelfogók elsődleges szerkezete meglehetősen hasonló: mindegyiknek
van egy jelvivőkötő (ligand binding domain, LBD; C’-végi E/F gomoly) és egy
DNS-kötő (DNA binding domain, DBD) gomolya; legtöbbjükben van még egy A/B és
egy D gomoly is.
A magjelfogók elsődleges szerkezete meglehetősen hasonló: mindegyiknek
van egy jelvivőkötő (ligand binding domain, LBD; C’-végi E/F gomoly) és egy
DNS-kötő (DNA binding domain, DBD) gomolya; legtöbbjükben van még egy A/B és
egy D gomoly is.
▪ Az N’-végi gomoly (A–B gomoly) nagyon változékony; hosszúságában és bázissorában is lényegesen eltér a különböző magjelfogókban. Ebben van egy DNS-kötő bázissor, amelyet AF1 (activation function-1) gomolynak nevezünk. Az AF1 a jelvivőtől függetlenül is szabályozza a génátírást, de a jelvivőt kötő gomoly is ellenőrzi. Ez a rész foszforileződhet, és a 3’-végi gomollyal kölcsönhatásban alakítja a fehérje térszerkezetét.
▪ A középső törzsökös DNS-kötő gomoly (C gomoly) meglehetősen egyforma mindegyik magjelfogóban. Két cinkujj mintázatot tartalmaz; az ujjakban 4 ciszteint köt össze a cinkion. Az egyikben (CI) P-doboz van; ez kötődik a fajlagos hormonkötő válaszelemhez, a másikban (CII) D-doboz van; ez a jelfogókettőst hozza létre.
▪ A jelvivőt kötő és a C gomolyt összekapcsoló kicsi kapocsrész a D gomoly.
▪ A C’-végi jelvivőkötő gomoly (E–F gomoly, LBD); ehhez kapcsolódik a társmagjelfogó is. 12 α-csavarodásból áll, térszerkezete nagyjából egyforma a magjelfogókban, egymástól a jelvivőkötő helyben térnek el. F felszíne az AF2 (activation function-2), amely az átírás fő irányítója. A G-fehérje jelfogókhoz hasonlóan működik: a jelvivővel kapcsolódva az AF2 kötődik a társszabályozókkal, ennek következtében válik hatékonnyá a jelfogó.
A szteroid jelvivők szokásosan a jelvivőkötő gomolyhoz kapcsolódnak, a zsírsav/foszfolipid alapú jelvivőknél inkább térszerkezeti szabályozás történik (pl. RORγ). Egyes magjelfogóknak van foszforilezési mintázata is, de a foszforilezési tevősítésük még nem tisztázott.