miRNS (mikro-RNS) micro RNA, miRNA (miRNA duplex) 20–25 nukleotidból álló, kétszálú törzsökös RNS; az egyik szál az irányítószál (guide stand, antisense stand) a másik a kísérőszál (passanger stand, sense stand). ~2000 féle miRNS ismert. Az miRNS gének kódolják, amelyek a genom 1–5%-át teszik ki. Előfordulnak a DNS egészén, még a fehérjét kódoló gének közteseiben, a nem átfordítódó szakaszaiban és jóval ritkábban a képezőkben is. Pl. az MCM7 fehérjét kódoló gén köztesében vagy a nem átfordítódó részében 3 miRNS gén csoportosul. Az miRNS-t kódoló géneket a polimeráz-II írja át.
Az miRNS két előalakból az elsődleges és az elő-miRNS*-ből formálódik:
• Elsődleges miRNS* (primary miRNA, pri-miRNA) nagy (legalább 1000 nukleotid hosszú). Ezt kódolja a gén.
Egy vagy több szárhurok van benne. Ezt ismeri fel a droshaegyütteshez* (Drosha Microprocessor complex), amely a drosha (RNáz-3) és a DGCR8 fehérje által kialakított ~600 kDa nagyságú együttes. A droshaegyüttes az elsődleges miRNS-t rögtön kapcsolja, és kivágja a szárhurkokat (~25 nukleotid) átlagosan 10 nukleotid hosszú véggel. Így keletkezik az elő-miRNS*.
A köztesekből származó elsődleges miRNS együtt fejeződik ki a gazda génnel, ezeket a droshaegyüttes még az elő-mRNS kivágása előtt átalakítja elő-miRNS-sé.
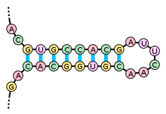 • Elő-miRNS (pre-miRNA, precursor miRNA)
kétszálú RNS jellegzetes szárhurokkal (stem-loop) és sok-A-as farkrésszel. Az
elő-miRNS-t a magból a kivivő fehérjepár* (nuclear
export machinery; exportin-5, RAN small DTPáz) a sejtplazmába szálltja,
ahol a DICER (RNáz-3, vég-RNáz), együttműködve a TRBP-vel, magához köti,
lehasítja a végi hurkot, kialakítva az érett miRNS-t, egyszerűen miRNS-t; ez az
AGO fehérjével kapcsolódik. Ebből jön létre a RISC, amelyben a miRNS
kísérőszála kilökődik, csak az irányítószál marad meg a kiegészítő mRNS
felismerésére. Az miRNS szabályozó hatása a RISC tagjaként érvényesül. (→RISC)
• Elő-miRNS (pre-miRNA, precursor miRNA)
kétszálú RNS jellegzetes szárhurokkal (stem-loop) és sok-A-as farkrésszel. Az
elő-miRNS-t a magból a kivivő fehérjepár* (nuclear
export machinery; exportin-5, RAN small DTPáz) a sejtplazmába szálltja,
ahol a DICER (RNáz-3, vég-RNáz), együttműködve a TRBP-vel, magához köti,
lehasítja a végi hurkot, kialakítva az érett miRNS-t, egyszerűen miRNS-t; ez az
AGO fehérjével kapcsolódik. Ebből jön létre a RISC, amelyben a miRNS
kísérőszála kilökődik, csak az irányítószál marad meg a kiegészítő mRNS
felismerésére. Az miRNS szabályozó hatása a RISC tagjaként érvényesül. (→RISC)
Az miRNS a génkifejeződés általános szabályozója; közbeavatkozó kis RNS. (→RNS-közbeavatkozás) A sejtmagban lévő miRNS csendesítheti vagy akár fokozhatja az átíródást azáltal, hogy kötődik az indítóhoz vagy a fokozóhoz, befolyásolja a DNS illetve a kromatin metilezését.
Sok miRISC van az energiatermecsben is. Nem tudjuk, hogy ezek az energiatermecs mRNS-eit is vagy csak a sejtmagból származó mRNS-eket támadják.
Az miRNS-nek testvérmásai (microRNA isoforms) is ismertek; ezek hosszukban, bázissorukban vagy mindkettőben és leginkább az 5’- vagy 3’-végen térnek el egymástól. Ennek alapján ötféle miRNS-eket különböztetünk meg:
▪ törzs miRNS-ek canonical microRNAs;
▪ 5’-végi testvérmás miRNS-ek 5′ isomiRs;
▪ 3’-végi testvérmás miRNS-ek 3′ isomiRs;
▪ sokalakú testvérmás miRNS-ek polymorphic isomiRs;
▪ vegyes testvérmás RNS-ek mixed type isomiRs.