sejtkör cell cycle a sejt osztódási folyamata, amelyben nyugalmi állapotából kettőződve kerül ismét nyugalmi állapotba.
Találatok címszavakban (5 szócikk):
DNS-kettőződés és a sejtkör A kettőződés az S-szakaszban megy végbe, 6–8 óra alatt. Előkészítése azonban már a sejtosztódás végén az M-szakaszban és a G1-szakaszban elkezdődik a kettőződési előössztes kialakulásával.
sejtkörellenőrző pontok cell cycle checkponint a sejtkör megállását vagy folytatását lehetővé tevő szakaszok. Ezekben ellenőrző fehérjék figyelik, hogy történt-e hiba, például a DNS-képzésben. Ha a bázisok nem megfelelők, a javítórendszerek kijavítják a hibát. A sejtkör csak akkor folytatódhat, ha a ellenőrzőpont mindent rendben talál.
Nagyon sok ellenőrző pont van; a három legjelentősebb: a G1-ellenőrző pont (R-pont, G1–S átmenet), a G2-ellenőrzőpont (G2–M átmenet) és a középszakaszi ellenőrző pont, amely az osztódási orsó (spindle apparatás) kialakulását ellenőrzi. Az osztódási orsó sejtvázi képződmény, a kromatídákat irányítja a sejt két szélére. A gyakorlatban mikor sejtellenőrző pontról beszélünk, ezeket értjük alatta.
Megkülönböztetnek továbbá biovegyi ellenőrző pontokat, amely a sejtkörben végbemenő biovegyi folyamatok sokaságának rendezői, a sejtkörrel összhangban dolgoznak, gátló (negatív) és serkentő (pozitív) szabályozók. Lehetővé teszik, hogy egymástól térben és/vagy időben elválasztott események kapcsolatba kerüljenek. Ezeket az ellenőrző pontokat két csoportba sorolják:
▪ Az első az önmaguktól nem működök; őket bekapcsolják, ha szükséges részvételük a sejtkörben, és kikapcsolják, ha már nem.
▪ A második csoportba tartozók önállóan tevősödnek, és akadályozzák a sejt előrehaladását. Ha a körülmények rendeződnek, kikapcsolódnak.
▪ G1–S átmenet. A G1-ellenörző (G1 phase cell cycle checkpoint) pont az első és a legjelentősebb; ez voltaképpen az R-pont, amely az átmenet elején helyezkedik; a sejt osztódásra való alkalmasságát (megfelelő a méret, a tápláltság, rendben van a jelzésközvetítés, hibátlan-e a DNS). A sejt az osztódási jelre csak a G1-szakaszának második kétharmadában, tehát korlátozott ideig képes válaszolni, és eldönteni, hogy véghezviszi-e a sejtkört. Ezt a korlátozott idejű szakaszrészt nevezik R-pontnak (restriction point [R point]); a sejtburjánzás meghatározó pontja). Ha a sejt nem folytatja a sejtkört, visszatér a nyugalmi állapotába. Ha a körülmények nem megfelelők a sejtosztódásra, a sejtkör megáll a G1–S átmenetben (G1-arrest).
G2-szakasz (G2–M átmenet). A G2-szakasz (G2 phase cell cycle checkpoint) az előkészület a sejt kettéválására, szükséges fehérjék (enzimek) képzésével, továbbá az S-szakaszt ellenőrzése, hogy hibátlan-e a keletkezett DNS, megtörtén-e teljesen a kettőződés. Ez a G2-ellenőrző pont feladata. Ha nincs rendben, feltartóztatja a folyamatot, amíg, hogy legyen idő kijavításra. Ha nem sikerül a javítás, a sejt a sejtvégzetbe megy.
▪Középszakaszi ellenörző pont* (metaphase chechkpoint) a sejtosztódás utolsó szakaszában (M-szakasz) azt ellenőrzi hogy minden egyes kromatida, meglelő hosszúságú mikrocsövecskével, tökéletesen kapcsolódjék a mozgóközéppel.
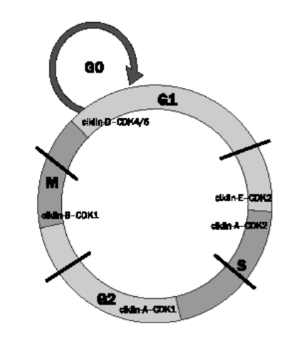 sejtkör felosztása Két fő szakasza: az S-szakasz, amelyben a DNS
megkettőződik, és az M-szakasz, a sejt kettéválásának, osztódásának a szakasz.
Mindegyik előtt van egy-egy átjárás és átmenet. Az S-szakasz előtt a G1-átjárás
és a G1–S átmenet, az M-szakasz előtt G2-átjárás és a G2–S
átmenet. Az átmenetek a lényegesek, ezek ellenőrző pontja (sejtköri ellenőrző
pontok) döntik el, hogy a sejt beléphet-e a főszakaszba. Az ábra vázlatosan
mutatja. (A G az angol gap
(szünet, kihagyás) szót, az S a synthesis, az M pedig a mitosis, meiosist
jelöli.)
sejtkör felosztása Két fő szakasza: az S-szakasz, amelyben a DNS
megkettőződik, és az M-szakasz, a sejt kettéválásának, osztódásának a szakasz.
Mindegyik előtt van egy-egy átjárás és átmenet. Az S-szakasz előtt a G1-átjárás
és a G1–S átmenet, az M-szakasz előtt G2-átjárás és a G2–S
átmenet. Az átmenetek a lényegesek, ezek ellenőrző pontja (sejtköri ellenőrző
pontok) döntik el, hogy a sejt beléphet-e a főszakaszba. Az ábra vázlatosan
mutatja. (A G az angol gap
(szünet, kihagyás) szót, az S a synthesis, az M pedig a mitosis, meiosist
jelöli.)
A nyugalmi állapot a sejt tevékenységének az ideje, ebben látja el a feladatát. Ezt a szakaszt G0-nak szokták nevezni. Valójában a nyugalomban lévő sejt eleve G1-ben van; ugyanis az osztódás után megszülető két új sejt (leánysejt) életét a G1 szakaszban kezdi – azaz rögtön ebbe kerül, ebben válik éretté (növekszik, növeli sejtszervecskéit, növeszti járulékos szerveit, például csillók stb.), és működik. Ha hosszabb ideig nem válaszol az osztódási jelekre, azaz már nem osztódik többet, de a feladatát ellátja, akkor kerül G0-ba. A G0-szakasz (quiescence, elcsitult) tehát olyan sejtet jelöl, amely megfelelően működik, de már nem osztódik, mert nem alkalmas a jel fogadására, például nincs jelfogója. Ilyenek az izom- és idegsejtjeink; felnőtt korban ezek már nem osztódnak egyáltalán. A nyugalmi sejt addig marad a G1-ban, ameddig nem kap engedélyt osztódásra, de az engedélyre való válasz is ahhoz kötött, hogy kellőképpen megérett-e már.
A szervezet osztódási jeleket küld a sejt közvetlen közelébe, ezeket osztódási/növekedési jeleknek hívunk. Ha a sejt már eltöltött megfelelő időt G1-ben, tud ezekre válaszolni. A kellő válaszhoz 10 óra szükséges. A G1–S átmenet elején lévő ellenőrző pont, az R-pont (restriktion point [akadályozó]), amely a G1–S átmenet kezdetén van, ellenőrzi, hogy a sejt megfelelő-e az osztódásra, és ha igen tovább engedi. Az R-pont azért van a G1-ben viszonylag távol, hogy a sejt növekedhessen kellőképpen, amíg eléri az R-pontot.
A G1 végén, a R-ponton átjutva minden az osztódásról szól, a sejt nem működik tovább, nem is igazán növekszik. Az R-ponton átlépve a G1–S átmenetben egy kis idő kell még a felkészülésre (enzimek előállítása) az osztódáshoz. Az S-szakaszba lépve, azonnal kezdődik a DNS-kettőződés. Ha ez befejeződött, a G2-átjárás feladata magára a sejtszétválásra történő előkészület. 12–13 óra alatt kettéválik a sejt.
sejtkör molekulatörténései A sejtkört alapvetően háromféle fehérje vezérli és szabályozza. Ezek a ciklinek, a CDK (ciklin dependen kinase) és a CDK gátlók (CDKI). A sejtkör alatt szakaszfüggően keletkeznek. A ciklin a CDK-vak kapcsolódik, csak együtt hatásosak. A CDK szerin/treonin fehérje-kináz, foszforilez, de csak azt a fehérjét, amelyet a ciklin kapcsol, önmagában hatástalan. A ciklin a szabályozó egység, ő választja ki a foszforilezendő fehérjét. A CDK gátlók akadályozó hatásukkal szabályoznak.
Természetesen más fehérjék is részt vesznek. A két legjelentősebb a CAK és az APC. Az CDK a ciklinhez kapcsolódva válik tevékennyé, de nem lesz teljes értékű. A megfelelő foszforilezéséhez szükséges, hogy a CAK a T-hurokra foszfátcsoportot helyezzen. A CDK tehát csak a ciklin és a CAK együttes hatására képes megfelelően foszforilezni.
Az APC egy E3-ubikvitin-ligáz, a sejtkör lebontandó fehérjéit jelöli (kapcsolódik hozzájuk), hogy a 26S fehérjebontacsban szétessenek. Fehérjebontással szabályoznak. Meghatározók, amikor a ciklin-B-ket kell hirtelen lebontani az M-szakaszban.
■ G1, G1–S átmenet A pRb a kulcsszereplő, kapcsolódik az E2F átírásfehérjéhez, ennek következtében a kettős áthelyeződik az E2F indítójának tevősítő részéről a gátló részére. Az E2F társul a kettősödési társával, a DP1, ez része a pRb–E2F együttesnek, amely a DNS-hez az E2F indítójánál kapcsolódik. Az pRb–E2F célgénjei elősegítik a G1–S átmenetet, de a DNS-kettőződését is. Néhány célgén: DNS-polimeráz, A-ciklin, timidin-kimáz, dihidrofolát-reduktáz, CDK1/2 stb.
A G1–S átmenet a pRb gátló hatásának megszüntetésével, a pRb foszforilezésével indul. A felszabadult E2F beindítja a célgéneket, az S-szakaszhoz szükséges enzimek termelését. Megtörténik az előkészület a DNS kettőzésre.
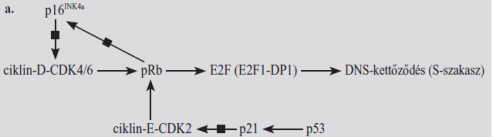 A pRb foszforilezését ciklin-D–CDK4 vagy CDK6 végzi a G1-szakasz
közepétől az R-pontig.
A pRb foszforilezését ciklin-D–CDK4 vagy CDK6 végzi a G1-szakasz
közepétől az R-pontig.
A ciklin-D–CDK4/6 a sejtkörnek ebben az időszakban keletkezik osztódás jelzések hatására, amelyeket sokféle jelút közvetít. Elsősorban a MAPK; innen cMYC, cFOS, cJUN átírásfehérjék szállítják az üzenetet a DNS-hez.
A folyamat fokozatos, és önmagát erősíti: a pRB folyamatosan válik el az E2F1–DP1 fehérjétől, egyre több gén íródik át, többek között a ciklin-E génje is. Az így keletkező ciklin-E–CDK2 pár is foszforilezi a pRb-t, ami további E2F1–DP1 felszabadulásához vezet, ez pedig még inkább serkenti a ciklin-E gént, még több ciklin-E–CDK2 keletkezik, tovább fokozva a pRB foszforilezését, főleg a G1-szakasz végén és a G1–S átmenetben. Voltaképp a ciklin-E–CDK2 irányítja a sejtet az S-szakaszba. Valószínű azonban, hogy a ciklin-E–CDK2 más módon is segíti az átmenetet az S-szakaszba. A CDK2–ciklin-E saját gátlóját, a p27Kip1 CDK-gátlót is foszforilezi, vagyis gátolja, így a hatása még kifejezettebb.(Ábrán nincs jelölve.)
Az R-ponttól jelennek meg ciklin-D–CDK4/6 gátlói, és elkezdődik a ciklin-D lebomlása. A p21 és a p16 a meghatározó CDK-gátló, az átmenet fő szabályozója. A p21 a p53 hatására képződik, akadályozza mind a három CDK-t (CDK2, -4 és -6). A p16 (az ábrán p16INK4a) gátolja a ciklin-D–CDK4/6 tevékenységét. A pRB foszforilezésére képződik, annak ellensúlyozására.
■ S-szakasz Az S-szakaszban a ciklin-E–CDK2 hatása még tart, de fokozatosan alábbhagy – a ciklin-E lebomlik –, helyét a ciklin-A–CDK2 veszi át, és irányit. A ciklin-E–CDK2 a kettőződéshez szükséges fehérjéket, enzimeket gyűjti össze. Ha elegendő az egyszeri kettőződéshez, a ciklin-A– CDK2 szintje emelkedi, és visszaszorítja a ciklin-E–CDK2 képződését. Ezzel, valamint a DNS-kettőződési rendszer egyik tagjának, a CDC6-nak a foszforilezésével akadályozza meg a kétszeres kettőződést.
A ciklin-A–CDK2 az S-szakaszban a sejtmagban van, előmozdítja a DNS-kettőződés indítását és véghezvitelét. A szakasz végén kapcsolódik a CDK1-gyel.
A DNS-másolódás időben szabályozott. Azt az időrendet, amelyben egyes DNS-részek másolódnak a nemzetközi irodalom replicatiom timingnak nevezi, magyarul osztódásidőzítés*. Törzsökös folyamat, szabályozatlansága betegséghez vezet. Elsősorban a kromatin állapota irányítja.
A DNS kettőződés végén két, testvér kromatidból álló kromoszóma keletkezik, tehát négy kromatida. Ezt nevezik tetradnak, magyarul négyes.
■ G2, G2–M átmenet. A ciklin-A–CDK1 és a ciklin-B–CDK2 irányítja; az utóbbi a G2-szakasz végén képződik, az osztódásba (M-szakaszba) lépés előtt fokozottan – ez indítja az osztódást olyan fehérjék foszforilezésével, amelyekre szükség van az M-szakaszban. A ciklin-A–CDK1 a késői S-szakasztól a G2-szakaszig marad meg, itt a ciklin-B veszi át a helyét.
A ciklin-A–CDK2, amely az S-szakaszban a sejtmagban van, a G2-ben bejut a sejtközpontba, és kötődik az osztódási orsóhoz. A részletek nem ismertek
■ M-szakasz (M- [mitosis] phase, mitozis) a sejt két sejtté
válásának az ideje: a sejt kettőzött DNS-állománya a sejtplazmával
együtt egyenlően szétosztódik két sejtbe; két azonos sejt keletkezik, amelyeket
leánysejteknek nevezünk. Lényegében két eseményből áll: a
kettőződött DNS egyenlő szétválásából, és a sejtplazma lefűződéséből
(cytokinesis). A ciklin-A–CDK1 és a ciklin-B–CDK1
teszi lehetővé az M-szakaszba lépést. A ciklin-B–CDK1 már az S-szakasz végén
megjelenik, a sejtmagban folyamatoson a sejtosztódás felé irányítja
sejtet. Mennyisége és tevékenysége állandó az osztódásig, ahol hirtelen
megszűnik a ciklin-B lebontása miatt. Ez szükséges ahhoz, hogy a sejt elhagyja
az M-szakaszt, újra a G1-szakaszba jusson. A ciklin-B–CDK1 hirtelen lebontását az APC (anaphase-promoting
complex) végzi, amely E3
ubikvitin-ligáz. Az enzim jelöli a lebontandó ciklin-B-ket hozzájuk kötődéssel;
a 26S fehérjebontacsban esnek szét.
Az M-szakaszt öt alszakaszra bontjuk.
 ● Előszakasz. Az előszakaszban (prophase,
● Előszakasz. Az előszakaszban (prophase, profázis) alapvetően
háromféle változás megy végbe:
▪ Kialakul a kétsarkú sejt és az osztódási orsó. Az S-szakaszban kettőződött sejtközép – amely körül csillagalakban, folyvást változó mikrcsövecskék vannak – a szakaszban a sejt két végére vándorol, kialakítva a kétsarkú sejtet. A továbbiakban ez a két pont határozza meg a sejtben végbemenő mozgásokat.
A sejtközépből kinyúló mikrocsövecskék közöl az ellentétes irányba nézők hozzák létre az osztódási orsót. Az osztódási hálózat tehát a sejtközpontokból elágazó mikrocsövecskék rendszere, amely meghatározza a sejtben végbemenő mozgások irányultságát.
▪ Tömörödnek a kromoszómák. A 46 kromoszóma elhelyezése a sejtben is rendkívüli, a megkettőződött kromoszómák, amelyek mindegyikéből két azonos kromatida lesz, tehát 92. A kromatidapárok egymással maradnak, a befűződéseknél kapcsolódnak össze. A 92 kromatida mozgatása sejtben csak úgy lehetséges, hogy a kromoszómák számottevően tömörönek, és jóval kisebbek lesznek. Valóban a kromatin nagyfokú tömörödése miatt vastagodnak a kromoszómák, annyira, hogy a középszakaszban már fénymikroszkóppal is jól láthatók; ezek a középszakaszi kromoszómák (metphase chromosoms). Természetesen a kromoszómák működése teljesen megszűnik. A magvacska bomlik, de a maghártya még ép.
▪ A sejtváz átalakulása. A sejtek állékonyságát biztosító áthidaló hosszú mikrotubulusok eltűnnek, miként a köztesfonalak is (intermedier filaments). A sejt változékonnyá alakul, lekerekedik és környezetével is lazul a kapcsolata. A plazmahálózat és a Golgi-rendszer is kezd szétesni.
● Előközépszakasz (prometaphase, prometafázis).
Kétféle meghatározó esemény időszakasza:
▪ A maghártya lebomlása. A maghártya belső faláz tapadó, laminból álló rácsrost lebomlik, és felszakad a maghártyatér, belőle hólyagcsák keletkeznek (maghártyahólyagcsák), továbbá eltűnnek az átjárások is. A magtér szabadon hozzáférhető: a mikrotubulusok belenőnek a kromoszómákhoz.
▪ A mjkrotubulusok kötődése a kromoszómákhoz. A testvérkromatídák együtt vannak, egymáshoz fekvő oldalukon a befűződés fogja össze őket. A kromatídák másik oldalán korongszerű fehérjeképződmény jön létre, a kinetokor, magyarul mozgóözépehhez kapcsolódnak mikrocsövecskék. Minden kromatidának saját kinetokorja van, a pároké ellenkező irányba néz. (→kinetokor)
● Középszakasz (metaphase, metafázis). Ebben kialakul az egyenlítői lemez
(equatorial/metaphase plate) a sejt közepén, benne a kromoszómákkal. Ez
viszonylag sok időt vesz igénybe: a kromoszómákat a mikrocsövecskék mozgatják
ide-oda (a mikrocsövecske rövidül, hosszabbodik) míg egészen pontosan nem
kerülnek a helyükre az egyenlítői síkban. A végén minden kromatida mindkét
sejtsaroktól tökéletesen egyenlő távolságra van. A teljesen egy síkba
rendeződött kromoszómákat tartalmazó képződményt középszakaszi lemeznek*
(metaphase plate) nevezzük. Ez biztosítja, hogy a kromatidapárok egyik tagja a
sejt egyik sarkába, a másik a másik sarkában kerül, a leánysejtekben pontosan
egyforma DNS lesz.
●
Utószakasz (anaphase, anafázis). Két egymást átfedő esemény játszódik
benne:
▪ A kromoszómák szétválása. A testvér kromatídák még összekapaszkodnak, fehérjeössztes tartja egybe őket a befűződésben, de a kohezin is össszetartozást hoz létre a kromatídák egész hosszában. Ezek a kapcsolatok hirtelen jelre megszűnnek, a kromatidák elválnak, s egyedileg mozognak a hozzájuk közelebbi sarkokhoz, a kinetokorral kapcsolódott mikrocsövecskék húzzák őket úgy, hogy tubulinkettősök képződve egyre rövidülnek.
▪ Menyúlik az osztódási orsó. Ez a sarki mikrocsövecskék további növekedésével jön létre, miközben a sejt közepén az egymás melletti sarki csövecskék kinezinnel összetapadnak.
● Végszakasz. A végszakaszban (telephase) a sejtosztódás befejező része. Újraképződik a maghártya, lazul a kromatin: a kromoszómák kezdenek kromatinkötegekké alakulni. A sejtplazma még egységes, de formálódik az osztódási barázda. A sejtközpont tovább irányítja mikrocsövecskéket.
A sejtplazma szétválása már az utószakaszban elindul: az osztódási orsó, kromoszómákat tartalmazó egyenlítői síkjának megfelelően, vagyis a sejt közepén, a sejtfelszínen barázda jelenik meg. Oka, hogy ezen a területen, a sejthártya alatt aktinfonalakból és rövid miozinkötegeből, összehúzódásra képes gyűrű képződik (összehúzódó gyűrű). Az aktin–miozin kölcsönhatás következtében a gyűrű folyamatosan kisebbedik, húzza a sejthártyát, és egyre inkább mélyül az osztódási barázda, végül a sejt szétválik (cytochinesis).
sejtkörszabályozás olyan ellenőrző folyamat, amely eldönti, hogy sejt osztódik-e vagy marad nyugalmi állapotában, változik-e vagy a sejtvégzet áldozatává válik. Meghatározó a szövetek helyreállításában; zavara a ráksejt jellegzetessége. A sejtkört a ciklinfüggő kináz (CDK) és ciklin kettősök, és ezek gátlói (CKI), valamint különböző átírásfehérjék szabályozzák. Ezek a sejtkör szakaszai szerint általában mások, rendszerint az adott szakaszban képződnek.
Részleges egyezések (1 szócikk):
cell cycle inhibitor →sejtkörgátló