ugrálat*
transposon, transposable element (transzpozon); (egyéb nevek: jumping
gene, selfish NDA, molecular parasite, controlling element) mozgékony
DNS-bázissor; képes áttevődni a DNS másik helyére. Egyesek másolódás nélkül,
mások másolódással helyeződnek át. Az előbbiek száma állandó, mert csak áthelyeződnek.
Az utóbbiak száma viszont nő, mert a másolódott bázissor ékelődik be a DNS
valamely részébe, az eredeti – amelyről a beékelődő bázissor másolódik –
megmarad az eredeti helyén. Az ugrálatok áthelyeződését különböző állapotváltoztató
(metilezési, kromatinátalakulási) folyamatok szabályozzák a korai ébrény
fejlődésétől kezdve. Szabályozásukat környezeti hatások (ártmányok, öregedés,
életmód stb.) befolyásolják. Az ugrálatoknak két csoportját, a kivágódóugrálat*
és a visszugrálat*, különböztetjük meg.
■ kivágódóugrálat* (DNA transposon, class/type II transposon) másolódás nélküli áthelyeződésű mozgékony DNS-szakasz; a szakasz kivágódik és beékelődik. 1000–40 000 bázispár nagyságú. Két formája az önálló és a hiányos kivágódóugrálat.
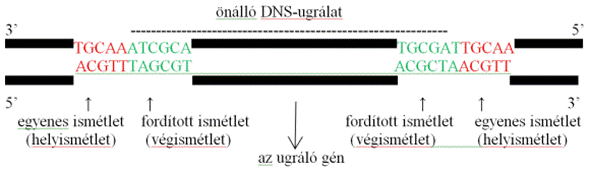 ▪ Önálló kivágódóugrálat*
tartalmazza a saját kivágását és áthelyezését végző enzimet (transzpozáz,
transposase) kódoló génrészt. A génrész négy képzőt tartalmazó olvasáskeret. Ez
közvetlenül kódolja a transzpozászt, RNS nem vesz részt benne. A génrész két
végén fordított ismétlet* (végismétlet*) van, az ugrálat ennek alapján
ismerhető fel. Az önálló kivágódóugrálat tehát a génrészből és a végismétletből
áll. A fordított ismétleteket egy-egy egyenes ismétlet* (helyismétlet*)
szegélyezi (target site duplication, TSD), ezek jelölik az ugrálat kivágásának,
illetve beékelődésének a helyét. A helyismétletek nem részei az ugrálatnak; az
ugrálatok kivágását követően szabályozhatják az utánuk lévő gének működését.
▪ Önálló kivágódóugrálat*
tartalmazza a saját kivágását és áthelyezését végző enzimet (transzpozáz,
transposase) kódoló génrészt. A génrész négy képzőt tartalmazó olvasáskeret. Ez
közvetlenül kódolja a transzpozászt, RNS nem vesz részt benne. A génrész két
végén fordított ismétlet* (végismétlet*) van, az ugrálat ennek alapján
ismerhető fel. Az önálló kivágódóugrálat tehát a génrészből és a végismétletből
áll. A fordított ismétleteket egy-egy egyenes ismétlet* (helyismétlet*)
szegélyezi (target site duplication, TSD), ezek jelölik az ugrálat kivágásának,
illetve beékelődésének a helyét. A helyismétletek nem részei az ugrálatnak; az
ugrálatok kivágását követően szabályozhatják az utánuk lévő gének működését.
▪ Hiányos kivágódóugrálat* csak külső enzim segítségével képes áthelyeződni, önállóan nem. Sok változata ismert, pl. van olyan, amely az önálló DNS-ugrálattól csupán néhány bázis törlődésében különbözik, de olyan is, amelyben az egész transzpozázt kódoló rész hiányzik stb.
Az önálló és a hiányos DNS-ugrálatok családot alkotnak. A DNS-ben több ilyen ugrálatcsalád van, ezeknek egymáshoz semmi közük.
■ visszugrálat (retrotransponson, class/type I transposon) másolódással áthelyeződő mozgékony DNS-szakasz. Áthelyezéséhez szükséges egy indító – innen kezdődik a fordított átírás –, valamint a fordított transzkriptáz, amely fordított átírással képezi a másolatot, továbbá egy RNSáz-H – ez bontja (szabadítja fel) az ugrálatot – és egy integráz, pl. a transzpozáz, amely beékeli az ugrálatot. Az önálló visszugrálatokban mindegyik tevékenység benne van. Háromfélét különböztetünk meg:
 ▪ hosszú végismétletű visszugrálat* (long terminal repeat, LTR) a végek hosszú (~300
bázispár) egyirányú ismétletei jellemzik (hosszú végismétlet*). Közéjük
tartoznak a belretrovírusok. Mindkét végismétletet egy-egy egyenes ismétlet
(helyismétlet*, direct repeat) szegélyezi. Ezek határozzák meg a beékelődés
helyét.
▪ hosszú végismétletű visszugrálat* (long terminal repeat, LTR) a végek hosszú (~300
bázispár) egyirányú ismétletei jellemzik (hosszú végismétlet*). Közéjük
tartoznak a belretrovírusok. Mindkét végismétletet egy-egy egyenes ismétlet
(helyismétlet*, direct repeat) szegélyezi. Ezek határozzák meg a beékelődés
helyét.
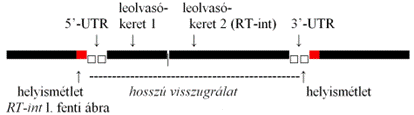
A gén részében 4 képező van:
Gag csoportfajlagos (group specific) antigént képez. Ez vírusszerű fehérje; tartalmazza a fordítottan átírt visszugrálat-DNS-t.
RT fordított transzkriptázt kódol, amely fordítottan átírt másolatot képez a visszugrálat-DNS-ről.
Prt proteázt kódol, amely lebontja a vírusszerű részecskét, ahogy a visszugrálat a sejtmagba kerül.
int integrázt kódol, ez szükséges az ugrálat beékelődéséhez.
Az utóbbi 3 részt (Prt, RT és int) magában foglaló DNS-szakaszt együttesen Pol-nak nevezzük. Az olvasáskeretek átfednek.
 ▪ hosszú elszórt visszugrálat* (long interspersed nuclear element, LINE) ősi DNS-elem, ~6 kb hosszú; a törzsfejlődés
során ékelődött be a DNS különböző helyeire. A genom ~15%-át teszi ki. Önálló,
nem LTR visszugrálat (non-LTR retrotransposon), enzimet kódol. Nincs végismétlete,
a génekhez hasonlóan, nem átfordítódó bázissorral kezdődik és végződik.
Indítója az 5’-UTR-ben van.
▪ hosszú elszórt visszugrálat* (long interspersed nuclear element, LINE) ősi DNS-elem, ~6 kb hosszú; a törzsfejlődés
során ékelődött be a DNS különböző helyeire. A genom ~15%-át teszi ki. Önálló,
nem LTR visszugrálat (non-LTR retrotransposon), enzimet kódol. Nincs végismétlete,
a génekhez hasonlóan, nem átfordítódó bázissorral kezdődik és végződik.
Indítója az 5’-UTR-ben van.
 ▪ rövid elszórt
visszugrálat* (short interspersed nuclear element, SINE) ~300 bázispárnyi,
nem önálló, nem LTR visszugrálat (non-LTR retrotransposon/retrosposon). Nem
kódol enzimet, csak segítséggel (pl. hosszú visszugrálat közreműködésével)
helyeződhet át – a hosszú visszugrálatból származik. Indítója az 5’-UTR-ben
van.
▪ rövid elszórt
visszugrálat* (short interspersed nuclear element, SINE) ~300 bázispárnyi,
nem önálló, nem LTR visszugrálat (non-LTR retrotransposon/retrosposon). Nem
kódol enzimet, csak segítséggel (pl. hosszú visszugrálat közreműködésével)
helyeződhet át – a hosszú visszugrálatból származik. Indítója az 5’-UTR-ben
van.
Az ugrálatok többsége „halott”: nem tevődik át; ezek másulás következtében elvesztik áthelyeződési képességüket. Jelentős részük epigenetikai szabályozókkal, pl. metilezéssel, a hisztonok módosításával némított, és csak nagyon kevés tevékeny, helyeződik át alkalomadtán. A kromatin tömörülése gátolja az áthelyeződést, ezért a heterokromatinban jóformán nincs is áthelyeződés.
Az emberi DNS ~50%-át ugrálatok teszik ki; összességében 2,5 billió bázispár nagyságúak. A gének 70–80%-ában vannak ugrálatok, zömében a köztesekben. Van olyan köztes, amely ~100 ugrálatot tartalmaz.
Az ugrálatok a DNS-állomány tartalékai, a DNS törzsfejlődési változásanak letéteményesei: Az áthelyeződés lehetővé teszi új gének kialakulását, pl. ha az ugrálat génbe ékelődik, megváltozik a gén szerkezete, az átalakulásból új gén keletkezhet. Így alakultak ki a törzsfejlődésben a környezeti változásokhoz alkalmazkodó gének. A másolódással áthelyeződők növelték a DNS-t, lehetővé téve, hogy a törzsfejlődésben bonyolultabb, a magasabban szerveződött élőlények létezéshez szükséges DNS-szerkezetek alakuljanak ki. Az ugrálatok szabályozhatják a gének működését, pl. áthelyeződésük valamely gén inditójának közelébe fokozhatja vagy gátolhatja az átíródást. Elősegítik a heterokromatin képződést, és ezzel rögzítik a DNS-t. A némított ugrálatok körüli kromatin ugyanis tömörül. Jelentősek az egyedfejlődésben a génműködés szabályozásával. Az ugrálat áthelyeződése azonban károsíthatja is a DNS működését: ha génbe ékelődik, megszűnik az adott gén működése, az indítórészhez kapcsolódása fokozhatja, pl. a daganatgének átíródását, de keletkezhet DNS-ingatagság, kromoszómatörés is. Ezek a DNS-hibák autoimmun, idegrendszeri, genetikai betegség és rák keletkezéséhez vezethetnek, fokozhatják a rákburjánzást.