kromatinszerveződés hagyományos szemlélet szerint a G0-sejtmagban a DNS és a kromatin szerveződésének három alapeleme fordul elő: 11 nm-es szál, 30 nm-es szál és a 300 nm-es szál.
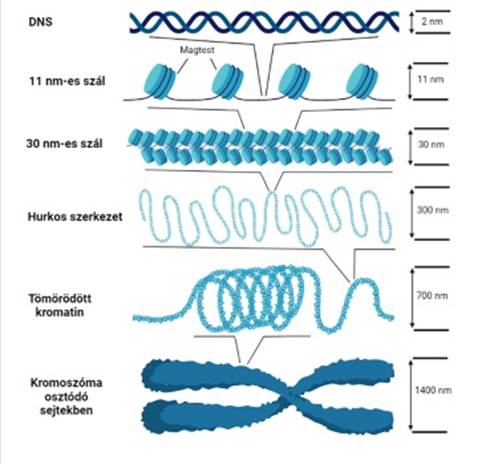 ▪ 11 nm-es
szál (11 nm fiber). A
magtestecsre feltekeredett DNS hosszú vékony szálat képez: ismétlődő
magtestecsek sorozatából és az azokat összekötő DNS-ből (linker DNA) áll. Gyöngyfüzérnek
(„beads-on-a-string”) is nevezik, mert hasonlít rá, de a fonál nem a gyöngyök
közepét átfúrva, hanem azokra feltekeredve halad. A 11 nm-es szál hossza
6–7-szer rövidebb, mint az eredeti DNS-é. A DNS és az összekötő hiszton (H1)
kapcsolata nem tisztázott; a legutóbbi elképzelés szerint a H1 hosszú C-végi
része kötődik a belépő és a kilépő összekötő DNS-sel, ~140 bázispárnyira a
magtestecstől; ez hozza létre a kromatin további szerveződését. A kapcsolódás
rugalmas, foszforilezésre érzékeny: könnyen felszakad, lehetővé téve a kromatin
megnyúlását, a DNS megközelítését.
▪ 11 nm-es
szál (11 nm fiber). A
magtestecsre feltekeredett DNS hosszú vékony szálat képez: ismétlődő
magtestecsek sorozatából és az azokat összekötő DNS-ből (linker DNA) áll. Gyöngyfüzérnek
(„beads-on-a-string”) is nevezik, mert hasonlít rá, de a fonál nem a gyöngyök
közepét átfúrva, hanem azokra feltekeredve halad. A 11 nm-es szál hossza
6–7-szer rövidebb, mint az eredeti DNS-é. A DNS és az összekötő hiszton (H1)
kapcsolata nem tisztázott; a legutóbbi elképzelés szerint a H1 hosszú C-végi
része kötődik a belépő és a kilépő összekötő DNS-sel, ~140 bázispárnyira a
magtestecstől; ez hozza létre a kromatin további szerveződését. A kapcsolódás
rugalmas, foszforilezésre érzékeny: könnyen felszakad, lehetővé téve a kromatin
megnyúlását, a DNS megközelítését.
▪ 30 nm-es szál (30 nm fiber, solenoid) sajátos ún. solenoid szerkezetűnek mondjuk, noha a solenoid szerkezetet sokféleképpen határozták meg, pl. az összekötő hisztonok egymással kötődve tengelyt képeznek, a magtestecsek ezen sorakoznak. Több ilyen képződmény tengelye tömörül úgy, hogy a magtestecsek többfelé helyezkednek el. De olvashatunk olyanról is, hogy a 10 nm-es szál láthatatlan tengely körül tekercselődik, egy-egy menetben ~6 magtestecs van.
▪ 300 nm-es kromatinszál ~100 kilobázisú hurkokat (hurokegységek*) alakító tömörödő kromatinszál. A hurkok részben a maghártyához, részben a magállományhoz rögzülnek. A további képződmények ennek tömörödéséből keletkeznek. (ábra: Baghy Kornélia)
Az új vizsgáló módszerekkel pontosabban tanulmányozható a kromatin szerveződése. A megfigyelések szemléletváltáshoz vezettek: megállapították többek közt, hogy a 30 nm-es szál nem alakul ki az élő sejtben; a kromatin változékony és szabálytalan formákat hoz létre a magtestecsek – főleg töltéshatásokra bekövetkező – mozgása következtében.
Kiderítették, hogy a DNS negatív töltése az összekötő szálakon megmarad, és az összekötő szál töltéskötésekkel kapcsolódhat az összekötő H1-hisztonhoz vagy más pozitív töltésű fehérjékhez, illetőleg pozitív ionokhoz. A maghisztonok pozitív töltései szintén lehetővé teszik a sokrétű kapcsolódásokat a DNS-sel, sőt távoli molekulákkal és a szomszédos magtestecsekkel is. A magtestecsek hisztonvégződéseinek összekapaszkodása a kromatintömörülés egyik alapfolyamata. Bizonyos fehérjék, pl. a kohezin, kifejezetten a magtestecsek társulását, vagyis a kromatin tömörülését okozzák. A kromatin szerveződését tehát a magtestecsek kapcsolódásai, mozgásai alakítják, amelyek alapvetően a magtestecseket körülvevő folyékony magállományban lévő molekulák töltésállapotától függnek.
Végeredményben a kromatin nagyon változékony formájú: a 10 nm-es kromatinszál igen sokféle alakzatba rendeződhet; hasonlóan a folyadékhoz – ezért nevezi a nemzetközi irodalom liquid-like chromatinnak. A kromatin környezetének töltésállapotát az ionok, kivált a magnéziumionok jelentősen befolyásolják.