indító* promoter a génátírást indító kétszálú DNS-szakasz. Megtalálható minden génben; általában a gén 5’-végének közelében van. Nem vesz részt az átírásban, csak irányítja: az átírásfehérje és az RNS-polimeráz kapcsolódik hozzá. Egy-egy indítóhoz több átírásfehérje is kötődhet. A fehérjéket folyvást képző gének indítói állandóan kapcsolják az RNS-polimerázt. Más gének indítói csak a hozzájuk kapcsolódó átírásfehérjék jelzésre.
Az indítók szerkezete alapjában különbözik attól függően, hogy melyik polimerázt indítják.
RNS-POLIMERÁZ-II INDÍTÓJA
A polimeráz-II általi átírását kezdő indító a fehérjéket kódoló gének indítója. Sajátos elemekből sokféleképpen tevődik össze; nincs mindegyik eleme jelen mindegyik génben. Az indító nagysága fajtánként eltérő, de az egyénekben is különböző; 100–1000 bázisnyi. Elemeit általánosan előforduló helyzetben a vázlatos ábra mutatja.
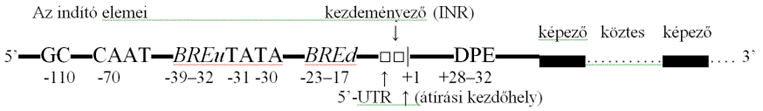
■ Átírási kezdőhely* (transcription start site, TSS) az indítómagban van; innen indul az átírás. Megfelel a gén azon bázisának, amelyről az elő-mRNS első nukleotidja másolódik, vagyis az először átíródó nukleotid helye. Az emberi sejtekben jóformán mindig két pirimidinbázist követő adenin. Ettől a bázistól számoljuk a gén bázisainak helyét; az átírási kezdőhely a +1 bázis. A 3’-vég felé lévő bázisok +, az 5’-vég felőliek - előjelet kapnak.
■ A fehérjeképző gének indítóinak két alapvető eleme a TATA-doboz és a kezdeményező* (INR), amelyek együttesen vagy külön találhatók meg. Mindkettő az indítómaghoz tartozik: önmagában képes indítani az átírást; és törzsökös: már az egysejtűekben is megtalálható. A TATA-doboz főleg a szövetfajlagos fehérjeképző gének indítója, az INR általános indító, a háztartási génekben van. Bonyolult fehérjéket (pl. MHC) kódoló gének indítóiban legtöbbször ez is, az is jelen van. Mindkettő köti a TFIId-t (átírásfehérje-IId), és alapot ad az indítóössztes kialakulására. (→génátírás)
• A TATA-doboz (5’-TATA-3’, TATA box) TATAWAW (TATAATAATR) alapbázissor (T = timin, A = adenin, R = purinbázis, W = ); nevét innen kapta. Első timinje az első átíródó bázistól (+1) ~30 bázispárnyira van az 5’-vég felé (-30). Az átírást meghatározott helyen, egy-két bázison belül indítja. A TFIIb-vel társult TFIId azonosítja, és legnagyobb összetevőjével, a TBP-vel (TATA binding protein) kapcsolódik hozzá. A TATA-doboz a gének ~10%-ban van jelen, és sokalakúan fordul elő. Bizonyos egynukleotidos változatai betegségre (pl. β-thalassemia) hajlamosítanak.
▪ A kezdeményező* INR (initiator element;
iniciátor) az átírási kezdőhelyet átívelő, YYA + 1 NWYY bázissor (az Y
pirimidinbázist, a W purinbázist az N bármelyik négy bázist jelöli), tehát a
génnek a kódoló részében is van (intragenic element). Nem behatároltan, hanem
legfeljebb 150 bázispárnyi sor valamely részén indít. Valószínű, hogy a gének
több mint felében jelen van. A TFIId kötődik hozzá, a TAF1 és a TAF2 alegység
ismeri fel.
■ BRE a TFIIb-t felismerő elem (B [TFIIB] recognition elements, BRE), a TATA-doboz egyik és másik oldalán helyezkedik el. Az 5’-vég felé lévő (BREu) -38–32 bázisnyira elhelyezkedő GCGCGACGCC bázissor. A 3’-vég felé lévő (BREd) -23–17 bázisnyira van: GATTATATGTGTGTG bázissor. Különleges, hogy a BRE fékezheti is az átírást.
■ A TATA-doboz nélküli génekben gyakran fordul elő a CAAT-doboz és a GC-doboz, de más elemek is:
▪ CAAT-doboz (CAAT box, CAT box) feltűnő GGTCCAATCT nukleotidsor az átírási kezdőhelyétől ~70 bázisnyira az 5’-vég felé. Törzsökös mintázat; különböző átírásfehérjék kapcsolódnak hozzá.
▪ GC-doboz GC box (ritkán GSG box) jól megkülönböztethető GGGCGG bázissor, amely cinkujjas átírásfehérjéket (SP1, MIGI, CREA Wilms-daganat stb.) köt. Az 5’-vég felé van, ~110 bázispárnyira az átírási kezdőhelytől. Erősíti a génátírást.
▪ TATA-szerű doboz (TATA-like box) TTTCAA bázissor, amely azonban nagyon változatos. Gyakran a polimeráz-III-t kötő indítókban található.
▪ MTE (motif ten element) szintén a TFIId-vel társul, a +18–27 helyen lévő, törzsökös CGCAGACGCGCAACGGC bázissor.
▪ TCT törzsbázissor az rRNS-t képző génekben van. Hasonlóan az INR-hez, magában foglalja a kezdőhelyet (-2–+6), de működése más: nem kapcsolja a TFIID-t. Részletek nem ismertek.
■ Az indítónak az átírási kezdőhelytől a 3’-vég felé is vannak bázissorai; ez DPR (downstream promoter region), alsó indítószakasz. Két elemét írják le:
• DPE (downstream [core] promoter element) törzsbázissor RGWYV ????(AGGATTCAG/C) +28–32 bázisnyira van az átírási kezdőhelyétől a 3’-vég felé. A TFIId felismeri, és kapcsolódik hozzá. Ez teszi lehetővé, hogy a TFIId áthidalja az átírási kezdőhelyet.
▪ DBE (TFIId binding element) a TFIId-kötő elem, három alelemmel (subelement [S] I–III; SI, SII, SIII): SI: CTTC, SII: CTGT; SIII: AGC a +6–11, a +16–21 és a +30–34 helyen. Működését kevéssé ismerjük.Viszonya DPE-vel sem tisztázott.
(A kitevőkben lévő bázisok felcserélődhetnek.)
Az indítókban gyakoriak a DNS-mintázatok (másodlagos DNS-szerkezetek, non-B-DNA structures), amelyek sajátos rajzolatok, az átírásfehérjék ezeket ismerik fel. Ilyenek az E-doboz, a guaninnégyes (D-quadruplex), az i-mintázat (I-motif), DNS-hármas (DNA-triplex), Z-DNS (Z-DNA) és a DNS-kereszt (DNA-cruciform).
alsó és felső indítósor downstream, upstream promoter az indítónak az átírási kezdőhelytől számított bázissorai; az 5’-vég felé van a felső, a 3’-vég felé az alsó szakasz.
indítómag core promoter az indítónak az indítóössztest létrehozó szakasza; elegendő az elő-mRNS képzésének elindítására. Az átírási kezdőhelyet és attól felfelé és lefelé terjedő 30–50 bázispárnyi DNS-szakaszt foglalja magában. Tartozéka a polimerázkötő hely, a TATA-doboz, a kezdeményező (INR) és a BRE, a DPE, a DBE és a DPRs. Az indítómag elemeinek mindegyike nincs mindegyik indítómagban jelen, nincs olyan elem vagy elem összeállás, amelyik mindegy indítómagban előfordulna. Az átíródás idején az indítómag szabad DNS-szálon van; tehát olyan DNS-szakaszon, amelyik nem a magtestecsre tekeredett. Az indítómag nemcsak a génátírás kezdője, hanem legfontosabb szabályozója is.
A gének 5’-végét részleteiben feltáró vizsgálómódszerrel (CAGE) tett megfigyelések szerint az indítómagnak (az indítónak) két formája van, függően az átírási kezdőhely elhelyezkedéstől:
▪ a szoros helyzetű (sharp vagy focused) indító, amelyben az átírási kezdőhely néhány bázison belül rögzült;
▪ a tág helyzetű (broad vagy dispersed) indító; ebben az átírási kezdőhely ~100 bázisnyi szakasznak bármely részén lehet.
Valószínű, hogy a kétféle indító különböző indítási folyamatot jelöl. Erre utal az is, hogy a szoros helyzetű indító általában tartalmazza a TATA-dobozt, és az indítómag más elemei is gyakrabban fordulnak elő, mint a tág helyzetű indítóban. Az utóbbi általában TATA-hiányos, sok benne a CpG-sziget (CG-gazdag). A tág helyzetűek az általános indítók, ilyenek vannak a háztartási génekben, míg a szoros helyzetűek sajátosak, pl. a szövetfajlagos gének vagy a fejlődésben résztvevő sajátos gének indítói. Ezek főleg a magasabb rendű fajokban találhatók, tehát a törzsfejlődés későbbi szakaszaiban jöttek létre.
A kétféle indítómag szabályozásában is eltér: másként válaszolnak szabályozási folyamatokra, mások a szabályozó molekulák is.
rejtett indító cryptic promoter DNS-kívüli szabályozással bénított, szokásosan tétlen indító. Genetikai vagy epigenetikai változás tevősítheti, az átíródásból kóros peptid keletkezik – gyakori rákokban. Feltételezhető, hogy így keletkezhetnek antisense és interfering szabályozó RNS-ek is.
RNS-POLIMERÁZ-I INDÍTÓJA
A polimeráz-I általi átírását kezdő indító az rRNS-eket képező gének indítója. Két GC-gazdag bázissorból állnak, amelyek az átírási kezdőhely két oldalán, -45 és +25 szakaszban vannak. Ezek önmagukban indítják az átírást. Egyes indítókban vannak még az átírást fokozó elemek az 5’-vég felé a -180 és -105 közötti részben.
RNS-POLIMERÁZ-III INDÍTÓJA
A polimeráz-III általi átírását kezdő indító a tRNS-eket és az snRNS-eket, valamint az 5S rRNS-t képező gének indítója. Ezek az indítók törzsökös bázissorokat tartalmaznak, amelyek a génekben vagy a gének előtt vannak. A polimeráz-III ezeket azonosítja. Sokféleképpen működnek.