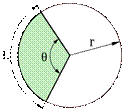
 kör, körív, körvonal zárt
görbével (önmagába visszatérő görbe vonal, körvonal) határolt síkalakzat,
melyben a görbe minden pontja a kör középpontjától egyenlő távolságban van. A
kör középpontja és kerülete közti terület a körlemez. A középpont és a
körkerület távolsága a kör sugara (radius, r). A kör két átellenes
pontját összekötő egyenes az átmérő (diameter, d), egyéb pontjait
összekötő egyenes pedig a húr. A körív a kör kerületének (a körvonalnak)
egy szakasza; nagyságát két sugár adja meg. A két sugár által közrefogott terület
a körcikk (az ábrán a zöld terület). A kör kerülete (k) 2π × r, ahol a π valós
szám, a kör kerületének és átmérőjének hányadosa = k / d; minden körnél azonos
érték. A körív egyenesen arányos a központi szögével, minél nagyobb a szög,
annál nagyobb a körív. A teljes szöghöz (360°) 2π, az egyenesszöghöz (180°) egy
π ívhossz tartozik. A központi szög (jele: θ)
csúcsa a kör középpontja, két befogója a kör sugara (r), az átfogója a körív
(L). A kört két ponton átszelő egyenes a szelő. A körrel egy ponton érintkező
egyenes az érintő. (Az ábrát Teleki Katalin készítette.)
kör, körív, körvonal zárt
görbével (önmagába visszatérő görbe vonal, körvonal) határolt síkalakzat,
melyben a görbe minden pontja a kör középpontjától egyenlő távolságban van. A
kör középpontja és kerülete közti terület a körlemez. A középpont és a
körkerület távolsága a kör sugara (radius, r). A kör két átellenes
pontját összekötő egyenes az átmérő (diameter, d), egyéb pontjait
összekötő egyenes pedig a húr. A körív a kör kerületének (a körvonalnak)
egy szakasza; nagyságát két sugár adja meg. A két sugár által közrefogott terület
a körcikk (az ábrán a zöld terület). A kör kerülete (k) 2π × r, ahol a π valós
szám, a kör kerületének és átmérőjének hányadosa = k / d; minden körnél azonos
érték. A körív egyenesen arányos a központi szögével, minél nagyobb a szög,
annál nagyobb a körív. A teljes szöghöz (360°) 2π, az egyenesszöghöz (180°) egy
π ívhossz tartozik. A központi szög (jele: θ)
csúcsa a kör középpontja, két befogója a kör sugara (r), az átfogója a körív
(L). A kört két ponton átszelő egyenes a szelő. A körrel egy ponton érintkező
egyenes az érintő. (Az ábrát Teleki Katalin készítette.)
Találatok címszavakban (8 szócikk):
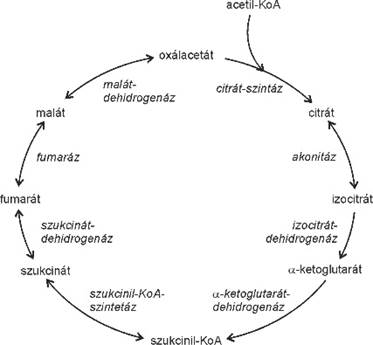 citrát-kör (Krebs-kör, Szent-Györgyi–Krebs-kör,
citromsavkör) tricarboxylic acid (TCA) cycle a sejtlégzés harmadik szakasza (→sejtlégzés),
energiatermelő körfolyamat, amely az anyagcsere-folyamatok sokaságában vesz
részt, az energiatermecs alapállományában megy végbe. Lévén körfolyamat, nincs
kezdete, sem vége; különböző molekulák más-más pontjain kapcsolódhatnak bele.
Talán az acetil-koenzim-A (acetil-KoA) társulás a leggyakoribb, amely a 4
szénatomos oxálacetáthoz (oxálecetsav) csatlakozik. A 2 szénatomos acetát
kötődik az oxálacetáthoz, és 6 szénatomos citrát keletkezik a citrát-szintetáz
közreműködésével; a KoA szabaddá válik, újrahasznosul.
citrát-kör (Krebs-kör, Szent-Györgyi–Krebs-kör,
citromsavkör) tricarboxylic acid (TCA) cycle a sejtlégzés harmadik szakasza (→sejtlégzés),
energiatermelő körfolyamat, amely az anyagcsere-folyamatok sokaságában vesz
részt, az energiatermecs alapállományában megy végbe. Lévén körfolyamat, nincs
kezdete, sem vége; különböző molekulák más-más pontjain kapcsolódhatnak bele.
Talán az acetil-koenzim-A (acetil-KoA) társulás a leggyakoribb, amely a 4
szénatomos oxálacetáthoz (oxálecetsav) csatlakozik. A 2 szénatomos acetát
kötődik az oxálacetáthoz, és 6 szénatomos citrát keletkezik a citrát-szintetáz
közreműködésével; a KoA szabaddá válik, újrahasznosul.
A következő lépésben a citrát elektronleadással átalakul (izocitráton keresztül) az 5 szénatomos α-ketoglutaráttá és CO2-vé; az elektront a NAD+ veszi fel, NADH +H+ keletkezik. Az α-ketoglutarátból a 4 szénatomos szukcinil-KoA képződik, KoA kötődésével. Ez szintén elektronleadással jár, itt is NADH + H+ jön létre és CO2 szabadul fel.
A további vegyületek a körben mind négy szénatomosak. A szukcinil-KoA szukcináttá alakul; a KoA leválik és energia szabadul fel, amely elegendő a GDP foszforilezéséhez, GTP-vé alakításához.
A szukcinátból két elektron leadásával fumarát lesz, az elektronokat a FAD veszi fel, FADH2 keletkezik.
A fumarátból malát lesz, a malátból pedig oxálacetát, és ezzel zárul a kör. A szukcinil-KoA – oxálacetát szakasz egyes lépései visszafordíthatók, a többi nem. A citrát-kör egésze megfordíthatatlan.
A citrát-kör végeredményben a piruvátból származó két szénatomos acetilcsoport bontása elektronelvonással úgy, hogy mind a két szénatom CO2-vé alakul; a szükséges oxigént vízmolekulából nyeri, a hidrogénjeit a NAD+ és a FAD veszi fel. A folyamat tiszta mérlege: az energiatermecs alapállományában lévő oxálacetáthoz több lépésben, köztes termékeken keresztül hozzáadjuk a következőket: acetil-KoA + 3 NAD+ + FAD + GDP + Pi. Visszanyerődik a KoA, keletkezik két molekula CO2, valamint 3 NADH+H+, 1 FADH2 és 1 GTP. A 3NADH+H+ körülbelül kilenc, a FADH2 pedig két ATP létrejöttéhez szükséges energiát hordoz.
A citrát-kör azonban nemcsak a sejtlégzésben vesz részt, hanem az anyagcsere-folyamatok zömében:
▪ Aminosavak lebontásában és újrahasznosításában cukorforrásként. 18-féle aminosavból keletkezhet glükóz, kizárólagosan a citrát-körön keresztül. Néhány (Ala, Gly, Ser, Cys, Thr, Trp) piruvátra bomlik, majd oxálacetáttá alakul. A Glu, Gln, Arg, His, Pro α-ketogluterátra, a Met, Ile, Val, Thr szukcinil-KoA-ra, az Asp, Asn, Phe, Tyr fumarátra vagy oxálacetátra (Asp, Asn) bomlik. Ezekből a molekulákból pedig szükség esetén megfelelő enzimek glükózt tudnak létrehozni. Ennek az éhezés állapotában van nagy jelentősége, mert ekkor a fehérjékből származó aminosavak a vér által a sejtekhez szállított megfelelő mennyiségű cukor forrásai.
A néhány aminosavból keletkező piruvát a glükóz bontásából keletkező piruváttal egyezően be is juthat a citrát-körbe, ahol az ismertetett módon energia szabadulhat fel belőlük.
▪ A zsírsavak energiájának kinyerésében. A zsírsavak lebontásából acetil-KoA keletkezik, ez pedig szintén a citrát-körön keresztül járul hozzá az ATP képződéséhez.
▪ Felépítő folyamatokban. Ezekben a citrát-körből kilépő molekulák vesznek részt, pl. a zsírsavak felépítésében a citrát, aminosavak keletkezésében az α-ketogluterát, a porfirinváz kialakításában a szukcinil-KoA.
DNS-kettőződés és a sejtkör A kettőződés az S-szakaszban megy végbe, 6–8 óra alatt. Előkészítése azonban már a sejtosztódás végén az M-szakaszban és a G1-szakaszban elkezdődik a kettőződési előössztes kialakulásával.
körforgás, keringés orbital angular momentum a test körpályán való mozgása; a test egy másik test (középtömeg*) körül mozog, például a Föld keringése a Nap körül. Olyan egyenes irányú mozgás, amely minden pillanatban változtatja az irányát, vagyis a középtömeggel szöget bezáró kerületi mozgás. Ugyanolyan erő hozza létre, mint az egyenes irányú mozgást. A keringő test azért nem halad egyenes pályán, mert egy másik test nem engedi, hogy eltávolodjék, például a tömegvonzás miatt. Ha megszűnik a középtömeghez rögzítő erő, a test a körpályára érintőleges egyenes mentén elrepül.
▪ A keringő test pályája kör. A középtömegtől a keringő testhez húzott sugár a vezérsugár; a test elmozdulásával a vezérsugár is elfordul. Azt a körívszakaszt, amelyet a test megtesz, ívnek nevezzük. Jele: i.
▪ A keringési idő egy kör megtételéhez szükséges idő. Jele: T. Mértékegységes: s (másodperc).
▪ A fordulatszám a keringő test által időegység alatt megtett körök száma. Jele: n. Mértékegysége. 1/s.
Egyenletes a körforgás, ha a test egységnyi idő alatt mindig egyforma nagyságú körívet tesz meg.
környezet surroundings (→rendszer, vegyfolyamat)
méhnyaktükör fénykibocsátó és nagyító rendszer, amelynek segítségével az élő szövet, a test egyes részeinek nagyított képét látjuk. A legtöbb kolposzkóp 5–25-szörös nagyításra képes, egyesek akár negyvenszeresre is, mások csak egyfélére; rendszerint tizenhatszorosra.
rendszer, környezet system, surroundings természettudományi fogalmak. A rendszer a világ (az anyagi valóság) vélt vagy valós határfelülettel elkülönített része, amelyet magunk választunk ki. Ilyen értelemben számtalan rendszer lehet, fizikai, vegyi és biológiai rendszerek sokasága. Például kémcsőben lévő folyadék, terem és tartozékai, vagy akár a Föld légköre, avagy gáz a léggömbben rendszer (a gáz eloszlásának, mozgásának és a léggömb falának a rendszere). Ami a rendszeren kívül van, az a környezet (surroundings). Vegyi rendszer például a vegyi anyagok keveréke, környezete az edényzet és minden más körülötte. Biológiai rendszer például a nyirokkeringés, az immunsejtek rendszere, környezete a szervezet többi része. Elemi biológiai rendszer a sejt; környezete a sejtkörüli állomány.
A rendszert állapotjellemzőkkel írjuk le, mint
hőmérséklet (T), nyomás (p), térfogat (V), anyagmennyiség (n) stb. Ennek
alapján lehet állandó nyomású (izobar), állandó hőmérsékletű (izoterm)
és állandó térfogatú (izochor).
A rendszer és a környezet kölcsönhatása szerint háromféle rendszert különböztetünk meg: elszigetelt, zárt és nyílt rendszert.
▪ Elszigetelt a rendszer, ha a határfelületén sem anyag, sem energia nem léphet át. Tehát a rendszerrel energiát sem közölhetünk, és nem is vonhatunk el.
▪ A zárt rendszer határfelületén energia átléphet, de anyag nem. Tehát energiát cserélhet a környezetével, például melegítjük. A zárt rendszer lehet állandó és változó térfogatú. Állandó például egy lezárt kémcső; térfogata nem változik. A dugattyús henger térfogata változik, függően a dugattyú állásától.
▪ A nyílt rendszer határfelületén az anyag is átléphet, nem csak az energia. Ilyen például egy folyadékot tartalmazó nyitott kémcső. Adhatunk hozzá vagy kiönthetünk belőle folyadékot. Melegítéssel pedig energiaváltozást idézünk elő.
A biológiai rendszerek nyílt rendszerek.
A
kölcsönhatást a rendszer fala határozza meg, például a hő szempontjából a fal
lehet hőszigetelő (adiabatikus) – ilyen az elszigetelt rendszer fala;
féligáteresztő (szemipermeábilis) vagy éppen hővezető.
A rendszereket csoportosíthatjuk a rendszer mérhető tulajdonságainak térbeli eloszlása szerint is:
▪ Egynemű rendszer homogeneous system (homogén
rendszer): mérhető tulajdonságai (állapotjellemzői) minden ponton
egyformák. Például egy jól elegyített oldat – hőmérséklete, nyomása stb. minden
részében azonos.
▪ Egyenetlen rendszer nonhomogeneous system (inhomogén
rendszer): mérhető tulajdonságai folyamatoson, lépésről lépésre változnak.
Például ha egy fémdarabot melegítünk, annak hőmérséklete folyamatosan nő, nem
ugrásszerűen változik.
▪ Egyveleg rendszer heterogeneous system (heterogén
rendszer): mérhető tulajdonságai ugrásszerűen változnak. Például olvadó
jég, amely az olvadása egy pontján folyadékká (víz) alakul, állapota
ugrásszerűen változik. Egyveleg, mert szilárd és folyékony összetevői is
vannak.
rendszerállapot* system state, thermodinamic state a rendszer mérhető fizikai tulajdonságainak összessége egy adott pillanatban. Függetlenül attól, hogy az állapot miként alakult ki.
rendszeregyensúly* system equilibrium a rendszert alkotó részecskék egyenletes eloszlása. Az állapothatározók egyike sem változik.
sejtkör cell cycle a sejt osztódási folyamata, amelyben nyugalmi állapotából kettőződve kerül ismét nyugalmi állapotba.
Találatok alcímszavakban (1 szócikk):
népességvizsgálat population study, population based study valamilyen csoport (pl. bennszülöttek, magyarok) vizsgálata; olyan személyeket vizsgál, akiknek fő jellegzetességei (kor, nem, betegség stb.) egyeznek. A jellegzetességek összegyűjtésével állapít meg irányvonalakat azonos állandó vagy változó mennyiségek szerint.
kiterjedt népességvizsgálat large scale population based study népességvizsgálat; valamely népesség zömét/egészét felölelő vizsgálat.
visszatekintő kiterjedt népességvizsgálat retrospective large scale population based study megtörtént adatok feldolgozásával a népesség zömében/egészében végzett vizsgálódás.
környezetvizsgálat ecological study (geographic study, correlation study) környezeti tényezőket vizsgál feltételezések bizonyítására, például a méhtestrák előfordulását veti össze a népesség táplálkozási szokásaival, keresve a kettő közötti összefüggést.
népességcsoport-vizsgálat population-based cohort study népességi csoportokat vizsgál; a személyeket együtt, nem pedig külön-külön.
visszatekintő népességcsoport-vizsgálat population-based retrospective cohort study már meglévő adatok népességi csoportok szerinti elemzése visszamenőleg.
szemléléses vizsgálat observational study (non interventional observational study) valamely népességben valamilyen módon kiválasztott személyeket tanulmányoz anélkül, hogy beavatkozna a sorsukba, például kezelné őket. Kiválaszt és megfigyel bizonyos betegségben szenvedőket, ez a betegségcsoport, és olyanokat, akiknek nincs az a betegségük. Az utóbbi az ellenőrző csoport, az ún. „egészségesek” csoportja. Több formája van:
elemző tanulmány analytic study a megfigyelés egyedekre terjed ki, a kiválasztott egyedek körülményeit tanulmányozza, például a kórok, kockázati tényezők stb. szempontjából.
esetsorozat vizsgálat case series study érdekes, különleges esetek tanulmányozása, számösszevetési értékelés nélkül.
keresztmetszeti vizsgálat cross sectional study egyfajta betegségben szenvedők különböző csoportjait vizsgálja a jelen helyzet szerint, például akiknél a betegség kezdődik vagy 5, 10 stb. éve fennáll.
leíró tanulmány descriptrive study a megfigyelés összesített adatokra (aggregated data) épül, például időskori méhtestrákok előfordulása. A vizsgálat népességi szintről tájékoztat, például összeveti a német és az angol népességben előforduló időskori méhtestrák eseteit.
valóhelyzet-vizsgálat real word study valamely készítmény, kezelés stb. vizsgálata válogatás nélküli népességben.
Részleges egyezések (11 szócikk):
bionomics →környezettan
cell cycle inhibitor →sejtkörgátló
enantiomer tükörképi (térközponti) azonmás (→téralakzati azonmások)
közepecsköri állomány Ez a sejtközpont másik összetevője. Tubulin és sok más (centrin, tektin, cenexin, nenein), köztük nem ismert fehérjéből áll. Ebben vannak a microcsövecske szervezési központok* (microtubule organization centers, MTOC); ezek képezik a mikrocsövecskéket. (→sejtközpont)
linea vonal ■ linea arcuata félkörvonal (→medencecsont/csípőcsont) ■ linea iliopectinea (→medencecsont) ■ linea terminalis határvonal (→medencecsont) ■ lineae transversae harántélek (→medencecsont/keresztcsont)
perivitelline space →sejthártyaköri térség
sejthártyaköri térség*MA
perivitelline space (perivitellinális térség) a megtermékenyítés
folyamán a petesejt hártyája és a fénylő burok között kialakuló térség neve. A
petesejt sejtplazmájába jutó ondósejt jeleket küld; ezek hatására a petesejt
kérgi szemcséi (cortical granules) kiürülnek a sejthártyaköri térségbe. A kérgi
szemcsékből felszabaduló enzimek úgy módosítják a fénylő burkot, hogy azon több
ondósejt nem tud áthatolni.
sejtkörellenőrző pontok cell cycle checkponint a sejtkör megállását vagy folytatását lehetővé tevő szakaszok. Ezekben ellenőrző fehérjék figyelik, hogy történt-e hiba, például a DNS-képzésben. Ha a bázisok nem megfelelők, a javítórendszerek kijavítják a hibát. A sejtkör csak akkor folytatódhat, ha a ellenőrzőpont mindent rendben talál.
Nagyon sok ellenőrző pont van; a három legjelentősebb: a G1-ellenőrző pont (R-pont, G1–S átmenet), a G2-ellenőrzőpont (G2–M átmenet) és a középszakaszi ellenőrző pont, amely az osztódási orsó (spindle apparatás) kialakulását ellenőrzi. Az osztódási orsó sejtvázi képződmény, a kromatídákat irányítja a sejt két szélére. A gyakorlatban mikor sejtellenőrző pontról beszélünk, ezeket értjük alatta.
Megkülönböztetnek továbbá biovegyi ellenőrző pontokat, amely a sejtkörben végbemenő biovegyi folyamatok sokaságának rendezői, a sejtkörrel összhangban dolgoznak, gátló (negatív) és serkentő (pozitív) szabályozók. Lehetővé teszik, hogy egymástól térben és/vagy időben elválasztott események kapcsolatba kerüljenek. Ezeket az ellenőrző pontokat két csoportba sorolják:
▪ Az első az önmaguktól nem működök; őket bekapcsolják, ha szükséges részvételük a sejtkörben, és kikapcsolják, ha már nem.
▪ A második csoportba tartozók önállóan tevősödnek, és akadályozzák a sejt előrehaladását. Ha a körülmények rendeződnek, kikapcsolódnak.
▪ G1–S átmenet. A G1-ellenörző (G1 phase cell cycle checkpoint) pont az első és a legjelentősebb; ez voltaképpen az R-pont, amely az átmenet elején helyezkedik; a sejt osztódásra való alkalmasságát (megfelelő a méret, a tápláltság, rendben van a jelzésközvetítés, hibátlan-e a DNS). A sejt az osztódási jelre csak a G1-szakaszának második kétharmadában, tehát korlátozott ideig képes válaszolni, és eldönteni, hogy véghezviszi-e a sejtkört. Ezt a korlátozott idejű szakaszrészt nevezik R-pontnak (restriction point [R point]); a sejtburjánzás meghatározó pontja). Ha a sejt nem folytatja a sejtkört, visszatér a nyugalmi állapotába. Ha a körülmények nem megfelelők a sejtosztódásra, a sejtkör megáll a G1–S átmenetben (G1-arrest).
G2-szakasz (G2–M átmenet). A G2-szakasz (G2 phase cell cycle checkpoint) az előkészület a sejt kettéválására, szükséges fehérjék (enzimek) képzésével, továbbá az S-szakaszt ellenőrzése, hogy hibátlan-e a keletkezett DNS, megtörtén-e teljesen a kettőződés. Ez a G2-ellenőrző pont feladata. Ha nincs rendben, feltartóztatja a folyamatot, amíg, hogy legyen idő kijavításra. Ha nem sikerül a javítás, a sejt a sejtvégzetbe megy.
▪Középszakaszi ellenörző pont* (metaphase chechkpoint) a sejtosztódás utolsó szakaszában (M-szakasz) azt ellenőrzi hogy minden egyes kromatida, meglelő hosszúságú mikrocsövecskével, tökéletesen kapcsolódjék a mozgóközéppel.
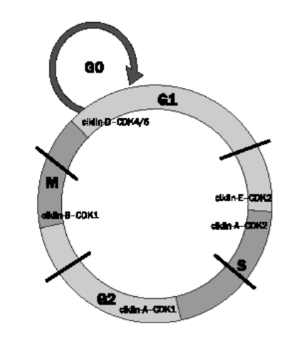 sejtkör felosztása Két fő szakasza: az S-szakasz, amelyben a DNS
megkettőződik, és az M-szakasz, a sejt kettéválásának, osztódásának a szakasz.
Mindegyik előtt van egy-egy átjárás és átmenet. Az S-szakasz előtt a G1-átjárás
és a G1–S átmenet, az M-szakasz előtt G2-átjárás és a G2–S
átmenet. Az átmenetek a lényegesek, ezek ellenőrző pontja (sejtköri ellenőrző
pontok) döntik el, hogy a sejt beléphet-e a főszakaszba. Az ábra vázlatosan
mutatja. (A G az angol gap
(szünet, kihagyás) szót, az S a synthesis, az M pedig a mitosis, meiosist
jelöli.)
sejtkör felosztása Két fő szakasza: az S-szakasz, amelyben a DNS
megkettőződik, és az M-szakasz, a sejt kettéválásának, osztódásának a szakasz.
Mindegyik előtt van egy-egy átjárás és átmenet. Az S-szakasz előtt a G1-átjárás
és a G1–S átmenet, az M-szakasz előtt G2-átjárás és a G2–S
átmenet. Az átmenetek a lényegesek, ezek ellenőrző pontja (sejtköri ellenőrző
pontok) döntik el, hogy a sejt beléphet-e a főszakaszba. Az ábra vázlatosan
mutatja. (A G az angol gap
(szünet, kihagyás) szót, az S a synthesis, az M pedig a mitosis, meiosist
jelöli.)
A nyugalmi állapot a sejt tevékenységének az ideje, ebben látja el a feladatát. Ezt a szakaszt G0-nak szokták nevezni. Valójában a nyugalomban lévő sejt eleve G1-ben van; ugyanis az osztódás után megszülető két új sejt (leánysejt) életét a G1 szakaszban kezdi – azaz rögtön ebbe kerül, ebben válik éretté (növekszik, növeli sejtszervecskéit, növeszti járulékos szerveit, például csillók stb.), és működik. Ha hosszabb ideig nem válaszol az osztódási jelekre, azaz már nem osztódik többet, de a feladatát ellátja, akkor kerül G0-ba. A G0-szakasz (quiescence, elcsitult) tehát olyan sejtet jelöl, amely megfelelően működik, de már nem osztódik, mert nem alkalmas a jel fogadására, például nincs jelfogója. Ilyenek az izom- és idegsejtjeink; felnőtt korban ezek már nem osztódnak egyáltalán. A nyugalmi sejt addig marad a G1-ban, ameddig nem kap engedélyt osztódásra, de az engedélyre való válasz is ahhoz kötött, hogy kellőképpen megérett-e már.
A szervezet osztódási jeleket küld a sejt közvetlen közelébe, ezeket osztódási/növekedési jeleknek hívunk. Ha a sejt már eltöltött megfelelő időt G1-ben, tud ezekre válaszolni. A kellő válaszhoz 10 óra szükséges. A G1–S átmenet elején lévő ellenőrző pont, az R-pont (restriktion point [akadályozó]), amely a G1–S átmenet kezdetén van, ellenőrzi, hogy a sejt megfelelő-e az osztódásra, és ha igen tovább engedi. Az R-pont azért van a G1-ben viszonylag távol, hogy a sejt növekedhessen kellőképpen, amíg eléri az R-pontot.
A G1 végén, a R-ponton átjutva minden az osztódásról szól, a sejt nem működik tovább, nem is igazán növekszik. Az R-ponton átlépve a G1–S átmenetben egy kis idő kell még a felkészülésre (enzimek előállítása) az osztódáshoz. Az S-szakaszba lépve, azonnal kezdődik a DNS-kettőződés. Ha ez befejeződött, a G2-átjárás feladata magára a sejtszétválásra történő előkészület. 12–13 óra alatt kettéválik a sejt.
sejtkör molekulatörténései A sejtkört alapvetően háromféle fehérje vezérli és szabályozza. Ezek a ciklinek, a CDK (ciklin dependen kinase) és a CDK gátlók (CDKI). A sejtkör alatt szakaszfüggően keletkeznek. A ciklin a CDK-vak kapcsolódik, csak együtt hatásosak. A CDK szerin/treonin fehérje-kináz, foszforilez, de csak azt a fehérjét, amelyet a ciklin kapcsol, önmagában hatástalan. A ciklin a szabályozó egység, ő választja ki a foszforilezendő fehérjét. A CDK gátlók akadályozó hatásukkal szabályoznak.
Természetesen más fehérjék is részt vesznek. A két legjelentősebb a CAK és az APC. Az CDK a ciklinhez kapcsolódva válik tevékennyé, de nem lesz teljes értékű. A megfelelő foszforilezéséhez szükséges, hogy a CAK a T-hurokra foszfátcsoportot helyezzen. A CDK tehát csak a ciklin és a CAK együttes hatására képes megfelelően foszforilezni.
Az APC egy E3-ubikvitin-ligáz, a sejtkör lebontandó fehérjéit jelöli (kapcsolódik hozzájuk), hogy a 26S fehérjebontacsban szétessenek. Fehérjebontással szabályoznak. Meghatározók, amikor a ciklin-B-ket kell hirtelen lebontani az M-szakaszban.
■ G1, G1–S átmenet A pRb a kulcsszereplő, kapcsolódik az E2F átírásfehérjéhez, ennek következtében a kettős áthelyeződik az E2F indítójának tevősítő részéről a gátló részére. Az E2F társul a kettősödési társával, a DP1, ez része a pRb–E2F együttesnek, amely a DNS-hez az E2F indítójánál kapcsolódik. Az pRb–E2F célgénjei elősegítik a G1–S átmenetet, de a DNS-kettőződését is. Néhány célgén: DNS-polimeráz, A-ciklin, timidin-kimáz, dihidrofolát-reduktáz, CDK1/2 stb.
A G1–S átmenet a pRb gátló hatásának megszüntetésével, a pRb foszforilezésével indul. A felszabadult E2F beindítja a célgéneket, az S-szakaszhoz szükséges enzimek termelését. Megtörténik az előkészület a DNS kettőzésre.
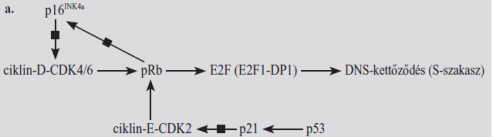 A pRb foszforilezését ciklin-D–CDK4 vagy CDK6 végzi a G1-szakasz
közepétől az R-pontig.
A pRb foszforilezését ciklin-D–CDK4 vagy CDK6 végzi a G1-szakasz
közepétől az R-pontig.
A ciklin-D–CDK4/6 a sejtkörnek ebben az időszakban keletkezik osztódás jelzések hatására, amelyeket sokféle jelút közvetít. Elsősorban a MAPK; innen cMYC, cFOS, cJUN átírásfehérjék szállítják az üzenetet a DNS-hez.
A folyamat fokozatos, és önmagát erősíti: a pRB folyamatosan válik el az E2F1–DP1 fehérjétől, egyre több gén íródik át, többek között a ciklin-E génje is. Az így keletkező ciklin-E–CDK2 pár is foszforilezi a pRb-t, ami további E2F1–DP1 felszabadulásához vezet, ez pedig még inkább serkenti a ciklin-E gént, még több ciklin-E–CDK2 keletkezik, tovább fokozva a pRB foszforilezését, főleg a G1-szakasz végén és a G1–S átmenetben. Voltaképp a ciklin-E–CDK2 irányítja a sejtet az S-szakaszba. Valószínű azonban, hogy a ciklin-E–CDK2 más módon is segíti az átmenetet az S-szakaszba. A CDK2–ciklin-E saját gátlóját, a p27Kip1 CDK-gátlót is foszforilezi, vagyis gátolja, így a hatása még kifejezettebb.(Ábrán nincs jelölve.)
Az R-ponttól jelennek meg ciklin-D–CDK4/6 gátlói, és elkezdődik a ciklin-D lebomlása. A p21 és a p16 a meghatározó CDK-gátló, az átmenet fő szabályozója. A p21 a p53 hatására képződik, akadályozza mind a három CDK-t (CDK2, -4 és -6). A p16 (az ábrán p16INK4a) gátolja a ciklin-D–CDK4/6 tevékenységét. A pRB foszforilezésére képződik, annak ellensúlyozására.
■ S-szakasz Az S-szakaszban a ciklin-E–CDK2 hatása még tart, de fokozatosan alábbhagy – a ciklin-E lebomlik –, helyét a ciklin-A–CDK2 veszi át, és irányit. A ciklin-E–CDK2 a kettőződéshez szükséges fehérjéket, enzimeket gyűjti össze. Ha elegendő az egyszeri kettőződéshez, a ciklin-A– CDK2 szintje emelkedi, és visszaszorítja a ciklin-E–CDK2 képződését. Ezzel, valamint a DNS-kettőződési rendszer egyik tagjának, a CDC6-nak a foszforilezésével akadályozza meg a kétszeres kettőződést.
A ciklin-A–CDK2 az S-szakaszban a sejtmagban van, előmozdítja a DNS-kettőződés indítását és véghezvitelét. A szakasz végén kapcsolódik a CDK1-gyel.
A DNS-másolódás időben szabályozott. Azt az időrendet, amelyben egyes DNS-részek másolódnak a nemzetközi irodalom replicatiom timingnak nevezi, magyarul osztódásidőzítés*. Törzsökös folyamat, szabályozatlansága betegséghez vezet. Elsősorban a kromatin állapota irányítja.
A DNS kettőződés végén két, testvér kromatidból álló kromoszóma keletkezik, tehát négy kromatida. Ezt nevezik tetradnak, magyarul négyes.
■ G2, G2–M átmenet. A ciklin-A–CDK1 és a ciklin-B–CDK2 irányítja; az utóbbi a G2-szakasz végén képződik, az osztódásba (M-szakaszba) lépés előtt fokozottan – ez indítja az osztódást olyan fehérjék foszforilezésével, amelyekre szükség van az M-szakaszban. A ciklin-A–CDK1 a késői S-szakasztól a G2-szakaszig marad meg, itt a ciklin-B veszi át a helyét.
A ciklin-A–CDK2, amely az S-szakaszban a sejtmagban van, a G2-ben bejut a sejtközpontba, és kötődik az osztódási orsóhoz. A részletek nem ismertek
■ M-szakasz (M- [mitosis] phase, mitozis) a sejt két sejtté
válásának az ideje: a sejt kettőzött DNS-állománya a sejtplazmával
együtt egyenlően szétosztódik két sejtbe; két azonos sejt keletkezik, amelyeket
leánysejteknek nevezünk. Lényegében két eseményből áll: a
kettőződött DNS egyenlő szétválásából, és a sejtplazma lefűződéséből
(cytokinesis). A ciklin-A–CDK1 és a ciklin-B–CDK1
teszi lehetővé az M-szakaszba lépést. A ciklin-B–CDK1 már az S-szakasz végén
megjelenik, a sejtmagban folyamatoson a sejtosztódás felé irányítja
sejtet. Mennyisége és tevékenysége állandó az osztódásig, ahol hirtelen
megszűnik a ciklin-B lebontása miatt. Ez szükséges ahhoz, hogy a sejt elhagyja
az M-szakaszt, újra a G1-szakaszba jusson. A ciklin-B–CDK1 hirtelen lebontását az APC (anaphase-promoting
complex) végzi, amely E3
ubikvitin-ligáz. Az enzim jelöli a lebontandó ciklin-B-ket hozzájuk kötődéssel;
a 26S fehérjebontacsban esnek szét.
Az M-szakaszt öt alszakaszra bontjuk.
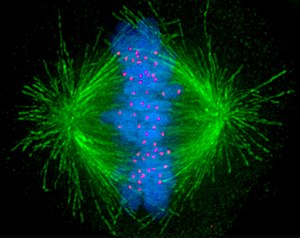 ● Előszakasz. Az előszakaszban (prophase,
● Előszakasz. Az előszakaszban (prophase, profázis) alapvetően
háromféle változás megy végbe:
▪ Kialakul a kétsarkú sejt és az osztódási orsó. Az S-szakaszban kettőződött sejtközép – amely körül csillagalakban, folyvást változó mikrcsövecskék vannak – a szakaszban a sejt két végére vándorol, kialakítva a kétsarkú sejtet. A továbbiakban ez a két pont határozza meg a sejtben végbemenő mozgásokat.
A sejtközépből kinyúló mikrocsövecskék közöl az ellentétes irányba nézők hozzák létre az osztódási orsót. Az osztódási hálózat tehát a sejtközpontokból elágazó mikrocsövecskék rendszere, amely meghatározza a sejtben végbemenő mozgások irányultságát.
▪ Tömörödnek a kromoszómák. A 46 kromoszóma elhelyezése a sejtben is rendkívüli, a megkettőződött kromoszómák, amelyek mindegyikéből két azonos kromatida lesz, tehát 92. A kromatidapárok egymással maradnak, a befűződéseknél kapcsolódnak össze. A 92 kromatida mozgatása sejtben csak úgy lehetséges, hogy a kromoszómák számottevően tömörönek, és jóval kisebbek lesznek. Valóban a kromatin nagyfokú tömörödése miatt vastagodnak a kromoszómák, annyira, hogy a középszakaszban már fénymikroszkóppal is jól láthatók; ezek a középszakaszi kromoszómák (metphase chromosoms). Természetesen a kromoszómák működése teljesen megszűnik. A magvacska bomlik, de a maghártya még ép.
▪ A sejtváz átalakulása. A sejtek állékonyságát biztosító áthidaló hosszú mikrotubulusok eltűnnek, miként a köztesfonalak is (intermedier filaments). A sejt változékonnyá alakul, lekerekedik és környezetével is lazul a kapcsolata. A plazmahálózat és a Golgi-rendszer is kezd szétesni.
● Előközépszakasz (prometaphase, prometafázis).
Kétféle meghatározó esemény időszakasza:
▪ A maghártya lebomlása. A maghártya belső faláz tapadó, laminból álló rácsrost lebomlik, és felszakad a maghártyatér, belőle hólyagcsák keletkeznek (maghártyahólyagcsák), továbbá eltűnnek az átjárások is. A magtér szabadon hozzáférhető: a mikrotubulusok belenőnek a kromoszómákhoz.
▪ A mjkrotubulusok kötődése a kromoszómákhoz. A testvérkromatídák együtt vannak, egymáshoz fekvő oldalukon a befűződés fogja össze őket. A kromatídák másik oldalán korongszerű fehérjeképződmény jön létre, a kinetokor, magyarul mozgóözépehhez kapcsolódnak mikrocsövecskék. Minden kromatidának saját kinetokorja van, a pároké ellenkező irányba néz. (→kinetokor)
● Középszakasz (metaphase, metafázis). Ebben kialakul az egyenlítői lemez
(equatorial/metaphase plate) a sejt közepén, benne a kromoszómákkal. Ez
viszonylag sok időt vesz igénybe: a kromoszómákat a mikrocsövecskék mozgatják
ide-oda (a mikrocsövecske rövidül, hosszabbodik) míg egészen pontosan nem
kerülnek a helyükre az egyenlítői síkban. A végén minden kromatida mindkét
sejtsaroktól tökéletesen egyenlő távolságra van. A teljesen egy síkba
rendeződött kromoszómákat tartalmazó képződményt középszakaszi lemeznek*
(metaphase plate) nevezzük. Ez biztosítja, hogy a kromatidapárok egyik tagja a
sejt egyik sarkába, a másik a másik sarkában kerül, a leánysejtekben pontosan
egyforma DNS lesz.
●
Utószakasz (anaphase, anafázis). Két egymást átfedő esemény játszódik
benne:
▪ A kromoszómák szétválása. A testvér kromatídák még összekapaszkodnak, fehérjeössztes tartja egybe őket a befűződésben, de a kohezin is össszetartozást hoz létre a kromatídák egész hosszában. Ezek a kapcsolatok hirtelen jelre megszűnnek, a kromatidák elválnak, s egyedileg mozognak a hozzájuk közelebbi sarkokhoz, a kinetokorral kapcsolódott mikrocsövecskék húzzák őket úgy, hogy tubulinkettősök képződve egyre rövidülnek.
▪ Menyúlik az osztódási orsó. Ez a sarki mikrocsövecskék további növekedésével jön létre, miközben a sejt közepén az egymás melletti sarki csövecskék kinezinnel összetapadnak.
● Végszakasz. A végszakaszban (telephase) a sejtosztódás befejező része. Újraképződik a maghártya, lazul a kromatin: a kromoszómák kezdenek kromatinkötegekké alakulni. A sejtplazma még egységes, de formálódik az osztódási barázda. A sejtközpont tovább irányítja mikrocsövecskéket.
A sejtplazma szétválása már az utószakaszban elindul: az osztódási orsó, kromoszómákat tartalmazó egyenlítői síkjának megfelelően, vagyis a sejt közepén, a sejtfelszínen barázda jelenik meg. Oka, hogy ezen a területen, a sejthártya alatt aktinfonalakból és rövid miozinkötegeből, összehúzódásra képes gyűrű képződik (összehúzódó gyűrű). Az aktin–miozin kölcsönhatás következtében a gyűrű folyamatosan kisebbedik, húzza a sejthártyát, és egyre inkább mélyül az osztódási barázda, végül a sejt szétválik (cytochinesis).
sejtkörszabályozás olyan ellenőrző folyamat, amely eldönti, hogy sejt osztódik-e vagy marad nyugalmi állapotában, változik-e vagy a sejtvégzet áldozatává válik. Meghatározó a szövetek helyreállításában; zavara a ráksejt jellegzetessége. A sejtkört a ciklinfüggő kináz (CDK) és ciklin kettősök, és ezek gátlói (CKI), valamint különböző átírásfehérjék szabályozzák. Ezek a sejtkör szakaszai szerint általában mások, rendszerint az adott szakaszban képződnek.