kettős ■ kettős kötés (→elektronkötés)
■ kettősödés dimerization (dimerizáció)
két azonos molekula összekapcsolódása egy vegyületté. ■ kettős vak vizsgálat double blind study (→klinikai vizsgálat)
Találatok címszavakban (5 szócikk):
kettőslény chimera, genetic chimerism (kiméra) olyan
lény, aki sejtjeiben kétféle DNS-állományt hordoz; mintha két lény lenne (két
lény egy testben).
Az elnevezés az ókori görög hitregékben lévő Kimérára utal, aki oroszlánfejű, kecsketestű, kígyófarkú tűzokádó szörnyeteg. Előfordul emberek között is, minden bizonnyal kivételes. Legtöbbjüknél soha nem derül ki a DNS-kettősség, esetleg csak orvosi vizsgálatkor. Másoknál előfordulhatnak eltérések, pl. a bőrfestékesedés eltérései (fokozott, csökkent, kevert), különböző színű szem, kétféle vércsoport, nemi szervi kettősség. Keletkezhet más egyén sejtjeinek a testbe jutásával, pl. anyai sejtek jutnak a magzatba, vagy fordítva: a magzat sejtjei az anyába, és ott megmaradnak (microchimerism). Avagy ikertestvérekben az egyik elhalásakor kerülnek annak sejtjei a másikba (twin chimerism). Előfordulhat véradáskor, valamint úgy is, hogy két idegen ondósejt termékenyít meg egy petesejtet, avagy kettőt, amelyek egyesülnek; egy személy keletkezik (tetragametic chimerism).
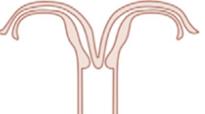 kettős méh didelphys uterus, duplex uterus a Müller-cső
rendellenességek harmadik csoportja, ritka (1:3000). A Müller-cső egyesülésének
elmaradásából származik. Két különálló méhüreg van, ezeket méhfal, vaskos
sövény választja el, az üregek nem közlekednek egymással. A méhnyak kettős, és
a legtöbb esetben hüvely sövény is van, amely lehet hosszanti, de haránt
elhelyezkedő is, annak minden következményével. Ha nincs elzáródás, lehet
tünetmentes, ha van, fájdalmak, alhasi kellemetlenség és/vagy szövetszaporulat,
fájdalmas havibaj és fájdalmas házasélet fordulhat elő. A felső húgyúti
rendellenességek gyakoriak, akárcsak a méhnyálkahártyásság.
kettős méh didelphys uterus, duplex uterus a Müller-cső
rendellenességek harmadik csoportja, ritka (1:3000). A Müller-cső egyesülésének
elmaradásából származik. Két különálló méhüreg van, ezeket méhfal, vaskos
sövény választja el, az üregek nem közlekednek egymással. A méhnyak kettős, és
a legtöbb esetben hüvely sövény is van, amely lehet hosszanti, de haránt
elhelyezkedő is, annak minden következményével. Ha nincs elzáródás, lehet
tünetmentes, ha van, fájdalmak, alhasi kellemetlenség és/vagy szövetszaporulat,
fájdalmas havibaj és fájdalmas házasélet fordulhat elő. A felső húgyúti
rendellenességek gyakoriak, akárcsak a méhnyálkahártyásság.
A kettős méhben fogant várandósság sokszor végződik vetéléssel, koraszüléssel, és nem ritka a magzat méhen belüli károsodása sem. Az MR mutatja a megváltozott anatómiát a legpontosabban. Műtéti megoldás legfeljebb többszöri vetélések után jön szóba, vagy ha kellemetlen panaszokat okoz.
petesejt–petedomb kettős oocyte-cumulus cell-complex, OCC a tüszőrepedéskor ez kerül ki az érett tüszőből, és jut a méhkürt rojtjai közé. A megtermékenyítés után a sugárkoszorú sejtjei azonban leválnak a fénylő burokról, és az ébrény, a fénylő burokba zárva vándorol át a méhkürt üregén a méhbe.
TSC-kettős tuberous sclerosis protein (TSC) 1/2 complex. A TSC1 (más néven: hamartin) és a TSC2 (más néven: tuberin) kettőst képezve hatékony, daganatgátló hatású, szabályozza az mTOR-jelközvetítést. Azonos nevű gének kódolják. A TSC2-ben GAP (GTPase activating protein) gomoly van; ez a RHEB-GTP-t RHEB-GDP-vé alakítja. A RHEB-GDP gátolja a mTORC1-t, kapcsolódik hozzá. A GTP-t kötő RHEB serkenti az mTORC1-et. A TSC1-ben nincs GAP gomoly, csupán rögzíti és védi a lebontástól a TSC2-t. (→gomoly, mTORC1, PI3K–jelközvetítés, RHEB)
XPC–RAD23b kettős egy 106 és egy 43 kDa tömegű fehérje rögzült társulása; a DNS-hibák nukleotidkivágó javításában vesz részt. A DNS-hibát ismeri fel, toborozza a TFIIh-t és az XPA-t. (→nukleotidkivágó javítás, RAD23b, XPC)
Találatok alcímszavakban (1 szócikk):
 foszfor (P) 15-ös rendszámú, 30,9738 atomtömegű, az
életfolyamatokhoz nélkülözhetetlen elem, a földkéreg tízedik leggyakoribb
eleme. Az élő szervezetben szerves vegyületekhez kapcsolódik, de előfordul
például kalcium-foszfát formájában is.
foszfor (P) 15-ös rendszámú, 30,9738 atomtömegű, az
életfolyamatokhoz nélkülözhetetlen elem, a földkéreg tízedik leggyakoribb
eleme. Az élő szervezetben szerves vegyületekhez kapcsolódik, de előfordul
például kalcium-foszfát formájában is.
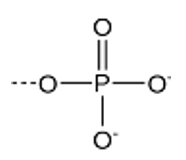
foszfátcsoport (PO42−) nevezik szervetlen foszfátnak is (inorganic phosphate), és Pi-vel jelöljük. Hasonlóan a foszforilcsoporthoz foszforsavszármazék.
kettős foszfát pirofoszfát anhidridkötéssel kapcsolt két foszfát; szervetlen foszfát, jelölése: PPi.
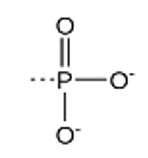 foszforilcsoport (PO32−) foszfort és oxigént
tartalmazó foszfátion, a foszforsav (H3PO4)
származéka; a szerves vegyületekben (aminosav,
glükóz, nukleozid) fordul elő. A biokémiában a szerves vegyületekben
lévő foszfort a szerves vegyület nevéhez írt P
betűvel jelöljük (ATP, P-szerin, G6P [glükóz-6-foszfát]).
foszforilcsoport (PO32−) foszfort és oxigént
tartalmazó foszfátion, a foszforsav (H3PO4)
származéka; a szerves vegyületekben (aminosav,
glükóz, nukleozid) fordul elő. A biokémiában a szerves vegyületekben
lévő foszfort a szerves vegyület nevéhez írt P
betűvel jelöljük (ATP, P-szerin, G6P [glükóz-6-foszfát]).
Részleges egyezések (5 szócikk):
dimerization domain kettősödési gomolyok bizonyos fehérjék azonos fehérjék való összekötődését elősegítő gomoly. (→fehérjegomoly)
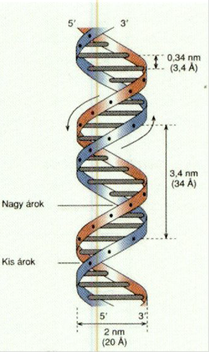 DNS kettős csavarodás* double helix A két csavarmenetű DNS-szál egymás
köré tekeredik ellentétes irányban (az egyik 5’→3’ a másik 3’→5’ helyzetű). Ezt
a szerkezetet nevezzük kettős csavarodásnak*; ez a kettős DNS-szál alakja. A
kettős szál átmérője 2 nm. A bázisok síkja a tengelyre merőleges, de a bázisok
egymáshoz viszonyítva 36o-kal elfordulnak, a köztük lévő távolság
0,34 nm. A csavarlat egy-egy 360º-os fordulata között tehát 10 nukleotid
található, a csavarulat távolsága pedig 3,4 nm. Az ettől eltérő tekeredés a
túltekeredés (supercoiling), amely lehet negatív és pozitív.
(→DNS-túltekeredés).
DNS kettős csavarodás* double helix A két csavarmenetű DNS-szál egymás
köré tekeredik ellentétes irányban (az egyik 5’→3’ a másik 3’→5’ helyzetű). Ezt
a szerkezetet nevezzük kettős csavarodásnak*; ez a kettős DNS-szál alakja. A
kettős szál átmérője 2 nm. A bázisok síkja a tengelyre merőleges, de a bázisok
egymáshoz viszonyítva 36o-kal elfordulnak, a köztük lévő távolság
0,34 nm. A csavarlat egy-egy 360º-os fordulata között tehát 10 nukleotid
található, a csavarulat távolsága pedig 3,4 nm. Az ettől eltérő tekeredés a
túltekeredés (supercoiling), amely lehet negatív és pozitív.
(→DNS-túltekeredés).
A két szálú DNS állékonyságában jelentős szerepe van a víztaszító hatásnak: A bázisoknak viszonylag nincs töltésük (víztaszítók), a foszfát pedig negatív töltésű (vízkedvelő), ezért kerül a foszfátot tartalmazó gerinc a DNS felszínére, és zárja be a bázisokat. A foszfát miatt a DNS felszíne negatív töltésű, és mert vizes közegben a töltéssel bíró (vízkedvelő) molekulák fordulnak a víz felé, a foszfát akadályozza, hogy a víz hozzáférjen az észterkötéshez, és felbontsa. A foszfátcsoport tehát védi a DNS-t a vízoldékonyságtól. A DNS szerkezetének állandóságában lényeges még az is, hogy a bázisok oxoformában vannak – ez teszi lehetővé, hogy a törzsfejlődésben bekövetkezhessenek szerkezetváltozások. Az enolforma kivételesen fordul elő. A kettős szál biztonságát még az is segíti, hogy a bázisok között jelentékeny London-féle kölcsönhatások is kialakulnak.
A DNS-szálak egymás köré tekeredése végbemehet jobbra és balra is, ezért jobbmenetes (a-, b-, és cDNS) és balmenetes (zDNS) csavarulat is keletkezhet. A bDNS-t (B-form DNA) tekintjük a DNS élettani formájának, a szervezetben általában ez fordul elő. Az aDNS és a cDNS környezeti hatásokra (alacsony nedvesség és sótartalom) a bDNS-ből keletkezik. Az aDNS-ben a bázispárok nem merőlegesek a kettős szál képzelt tengelyére – attól 19o-kal eltérnek –, aminek következtében a kis árok szinte eltűnik, a DNS megrövidül. Egy-egy menetnek megfelelően nem 10, hanem 11 nukleotid helyezkedik el. A cDNS a bDNS-től abban tér el, hogy benne egy-egy csavarulatot 9 nukleotid képez.
A zDNS nevét cikkcakkos (zig-zag) felcsavarodásáról kapta, amely a nukleozidok térbeli elhelyezkedéséből keletkezik. Szemben az anti helyzetű a- és bDNS-sel, a zDNS-ben a pirimidin- anti, a purinnukleozidok szün helyzetűek, ezért bennük csak egyféle árok található. Egy-egy csavarmenetet 12 nukleotid képez. A zDNS hosszabb és vékonyabb, mint a bDNS. Előfordul, hogy a DNS-nek csupán egy-egy rövidebb szakaszán alakul ki.
A két szálú DNS jóval állékonyabb, mint az egyszálú, mert a kettős csavarodás védi a bázisokat a károsító, másulást okozó vegyi és enzimhatásoktól. Erre utal pl. az is, hogy a citidin deaminacioja, aminek következtében a citidinből uracil keletkezik, csak az egyszálú DNS-en következik be, a kétszálun nem. További előnye a kettős szerkezetnek, hogy ugyanazt a genetikai üzenetet a sejt két szálon is tárolja; a szálak egymás kiegészítői. Ha hiba keletkezik, a nukleotidsor a másik szálról teljesen helyreállítható.
A kettős szerkezet rugalmas, a DNS-folyamatokban, pl. génátíródás, DNS-másolódás vagy DNS-javítás, a két szál szétválik, a folyamatokban résztvevő fehérjék csak így férhetnek a bázisokhoz. A szálak szétválasztását a a DNS-helikázok végzik a hidrogénkötések felbontásával ATP-energia segítségével.
ellenanyag–gyógyszer kettősök* antibody–drug conjugates Olyan kezelési mód, amelyben ráksejtre fajlagos ellenanyaghoz kapcsolják a gyógyszert, így az csak a ráksejtekhez kötődik. Előnye: a ráksejtekhez nagyobb töménységben kerül a gyógyszer, hatásosabban pusztít, az ép sejtekhez viszont alig kötődik, jóval enyhébb és kevesebb a mellékhatás. Továbbá hat a nem vagy lassan osztódó sejtekre is, szemben a sejtmérgező szerekkel, ezek ugyanis leginkább a gyorsan osztódó sejteket pusztítják.
kiméra (chimera) →kettőslény
nevezék valamely szakma vagy tudomány valamijét azonosító szó vagy kifejezés; a szakmát, tudományt művelők által adott elnevezés; lehet szó vagy szószerkezet, és társulhat tartozékelemekkel. (műszó, szakszó, szakkifejezés). (→fogalom)
nevezéktan nomenclature valamely tudomány nevezékeinek nemzetközi rendszere. Magában foglalja a névadást és a megnevezés rendszerét, az elnevezések értelmezését és rendszerezését.
kettős nevezéktan binomial nomenclature az élő szervezetek tudományos rendszertana, amelyben minden fajnak kettős neve van. Az első tag annak a nemzetségnek (állatoknál nemnek) a neve, amelybe a faj tartozik. A második tag a fajt a nemzetség többi tagjától megkülönböztető név. A nemzetségnevet tulajdonnévnek tekintjük, de csak az első tagot (a nemzetségnevet) írjuk nagy kezdőbetűvel; ezt gyakran rövidítjük (Streptococcus pneumoniae [S. pneumoniae]). A rendszert Carl von Linné (Carolus Linnaeus) alkotta meg az 1700-as évek közepén megjelent Species plantarum című munkájában (Linné-féle kettős nevezéktan). (→rendszertan).