retrovírus retrovirus fehérjékkel burkolt RNS-ből áll, amelyet lipidtok zár magába. Gömb alakú RNS-vírus, átlagos mérete 100 nm. Csak élő sejtben szaporodik, a sejtplazmában a vírus az RNS-ről a saját átíró enzimével (reverse trascriptase, fonák transzkriptáz*) kétszálú DNS-t képez, azaz a genetikai üzenet fordított irányú: RNS–DNS, nem pedig a szokásos DNS–RNS keletkezésű. Nevét a vírusok között a csak rájuk jellemző reverse trascriptase enzimről kapta: eleinte retravírusoknak nevezték ezeket a vírusokat, a mai neve elírás: retrovírus, s később ez honosodott meg a szakirodalomban is.
A retrovírus jellegzetessége, hogy a vírus-DNS, vagyis az RNS-ről keletkezett kétszálú DNS (provírusnak nevezzük) beékelődik a sejt DNS-ébe a vírus által kódolt integráz segítségével, és a sejt-DNS másolódásával együtt másolódik; a sejt egész életében megmarad. A provírust a sejt-DNS sajátjának tekinti, és szokványosan átírja; új vírusok képződéséhez szükséges fehérjék keletkeznek, a vírusok sokszorozódnak.
▪ A vírus szerkezeti fehérjéi. A retrovírus fő fehérjéje a GAG többfehérje (Group-specific antigen protein), amely kis fehérjékből és a kapcsoló részekből tevődik össze. A vírus GAG génje kódolja egyetlen fehérjeként; érésekor a proteáz darabolja kis fehérjékre, amelyek elsősorban a vírusok szerkezeti fehérjéit alkotják.
A GAG fehérje összetétele a retrovírusok fajtája szerint jelentősen eltérő, három törzsökös összetevője azonban mindegyikben fellelhető. Ezek az álladékfehérje* (matrix [MA] protein), a tokfehérje* (capsid [CA] protein) és a magtokfehérje* (nucleocapsid [NC] protein). Többes fehérjeként nem fertőző vírusszerű részecske (virus-like particle) előállítására képes. A proteáz által hasítva felszabaduló kis fehérjék hozzák létre a fertőző viriont. Az álladékfehérje a sejthártyához kötődve segíti a virion sejtfelszínre jutását, továbbá az ENV szénhidrátfehérjék beépülését. A tokfehérjék az RNS tokját alkotják. A magtokfehérje az RNS-hez kapcsolódik, lehetővé téve, hogy tok épüljön köré.
Az ENV (envelope) gén által kódolt fehérje (ENV fehérje) tüskefehérje (spike protein), a virion felszínéből tüskeszerű kinövéseket tartalmazó fehérje. A proteáz hasítja két összetevőjére, lehetővé téve, hogy az egyik kötődjék a sejtfelszíni jelfogóhoz, a másik pedig társjelfogóhoz (coreceptor). Az utóbbi közelíti a vírusburkot a sejthártyához, indítja a kettő összeolvadását. A tüskefehérjék száma vírusfajtaként tág határok között változik.
A retrovírusok a fenti szerkezeti fehérjék mellet egy vagy több szabályozó fehérjét is tartalmaznak, amelyek a vírusszaporodást és/vagy pl. a daganatkeletkezést szabályozzák.
▪ A vírus enzimei. A retrovírus életfolyamatához 3 enzim szükséges: a többfehérjéket hasító proteáz – ezt a PRO (proteáz) gén képezi; az RNS–DNS átalakítást végző fordított transzkriptáz, valamint az integráz, amely a vírus-DNS-t beékeli a sejt DNS-ébe. A fordított transzkriptázt és az integrázt a POL (polimeráz) gén kódolja.
▪ A vírus RNS-e. 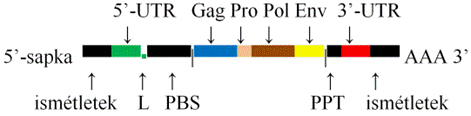 A vírus-RNS kettős RNS 5’-végi sapkával és 3’-végi
AAA-farokkal, hasonlóan az mRNS-hez, valamint nem átíródó bázissorokkal (UTR,
untranslated region). A közbenső rész kódolja a vírusfehérjéket. Az 5’-vég négy
elemet tartalmaz: Az 5’-végi sapka mellett rövid ismétletek vannak, ezek
szabályozzák a bázissorrendet a visszátíródásban. Ezt követi az 5’-UTR,
mellette az indítószál kötőhelye (PBS; primer binding site). Az 5’-UTR-ben van a nagyon rövid
jelző bázissor (L); az RNS becsomagolásához adja a jelzést. A 3’-végen szintén
négy elem, a sokpurinos szakasz (PPT [polypurin tract]), mellette a 3’-UTR van,
amelyet rövid ismétletek követnek, a legvége AAA-farok. A 3’-UTR adja a
jelzést az átíródásra. A közbenső részen négy törzsökös gén (Gag [group-specific
antigen gen], Pro [protease gene], Pol [polymerase gene] és Env
[envelope gene]) van; a retrovírusok fajtái szerint továbbiak is
előfordulnak.
A vírus-RNS kettős RNS 5’-végi sapkával és 3’-végi
AAA-farokkal, hasonlóan az mRNS-hez, valamint nem átíródó bázissorokkal (UTR,
untranslated region). A közbenső rész kódolja a vírusfehérjéket. Az 5’-vég négy
elemet tartalmaz: Az 5’-végi sapka mellett rövid ismétletek vannak, ezek
szabályozzák a bázissorrendet a visszátíródásban. Ezt követi az 5’-UTR,
mellette az indítószál kötőhelye (PBS; primer binding site). Az 5’-UTR-ben van a nagyon rövid
jelző bázissor (L); az RNS becsomagolásához adja a jelzést. A 3’-végen szintén
négy elem, a sokpurinos szakasz (PPT [polypurin tract]), mellette a 3’-UTR van,
amelyet rövid ismétletek követnek, a legvége AAA-farok. A 3’-UTR adja a
jelzést az átíródásra. A közbenső részen négy törzsökös gén (Gag [group-specific
antigen gen], Pro [protease gene], Pol [polymerase gene] és Env
[envelope gene]) van; a retrovírusok fajtái szerint továbbiak is
előfordulnak.
▪ RNS–DNS átalakulás. Az átalakulás bonyolult, soklépcsős folyamat, alapvetően a fordított transzkriptáz végzi. Egyéb fehérjék is közreműködhetnek, pl. a maghéjfehérje serkenti a folyamatot. A sejt egyik tRNS-se az elsődleges kötőhelyhez (PBS) kapcsolódik. A vírus-RNS a tRNS-től másolódik az 5’-végig, rövid DNS–RNS kettős jön létre. A másolás tehát nem az RNS-szál végén kezdődik. Az 5’-végi rövid DNS–RNS kettősből a fordított transzkriptáz eltávolítja az RNS-t, a DNS pedig áthelyeződik a vírus-RNS 3’-végére, a 3’-végi ismétletekhez kötődik. A fordított transzkriptáz ezt hosszabbítja meg, az RNS-t mintafelületként használva. Vagyis DNS–RNS kettős keletkezik. Ennek az RNS-szála egy rövid szakasz kivételével lebomlik. A másik DNS-szál ettől a rövid RNS-száltól másolódik a 3’-végig a sejt DNS-polimeráza által. A második DNS-szál leválik, és újra kereszteződik az 5’-végen lévő elsődleges kötőhellyel. Ez másolódik mindkét irányban úgy, hogy a vírus-RNS-nek megfelelő kétszálú DNS keletkezik (vírus-DNS). Az integráz ezt építi be a sejt DNS-ébe.
 ▪ Vírus-DNS. A 3’-UTR és az 5’-UTR is
kettőződik, ezért a vírus-DNS mindkét végén hosszabb, mint a vírus-RNS. A
végeken lévő UTR-ekből és ismétletekből alakul ki a hosszú végismétlet (long
terminal repeats), amely jellegzetes a vírus-RNS-ből keletkezett DNS-re.
▪ Vírus-DNS. A 3’-UTR és az 5’-UTR is
kettőződik, ezért a vírus-DNS mindkét végén hosszabb, mint a vírus-RNS. A
végeken lévő UTR-ekből és ismétletekből alakul ki a hosszú végismétlet (long
terminal repeats), amely jellegzetes a vírus-RNS-ből keletkezett DNS-re.
A vírus életfolyamata: fajlagosan kötődik jelfogóhoz, amely a sejthártyához kapcsolja; a vírusburok és a sejthártya összeolvad úgy, hogy a vírusmag a sejtplazmába kerül. A plazmában a fonák transzkriptáz a vírus-RNS-ből egyszálú DNS-t készít; ezt alakítja a sejt DNS-polimeráza kétszálú DNS-sé, amelyhez az integráz kapcsolódik. A fonák transzkriptáznak nincs hibajavító képessége, ezért sokszor keletkezik hibás vírus-DNS-t tartalmazó vírus. Csak néhány retrovírus képes a vírus-DNS-t bejuttatni a sejtmagba, a legtöbbnél a sejtosztódáskor kerül a vírus-DNS kapcsolatba a sejt-DNS-sel, amelybe az integráz beépíti. A polimeráz-II írja át, számos vírus-RNS keletkezik. A vírus összeszerelése legtöbbször a sejthártya belső felszínén megy végbe úgy, hogy a teljes vírus, a virion kiszabadul a sejtből.
▪ virion fehérjetokba (capsid, beltok*) csomagolt RNS-ből (vírusmag), a vírusmag körüli álladékból (matrix) és az egészet körbevevő kettős lipidburokból áll, amelybe a vírus által kódolt fehérjék is beépülnek. A kettős lipidburok a sejtfalból keletkezik, amikor a vírus kilép a sejtből (endosomal membrane trafficking). A burok felszínéből glikofehérje tüskék állnak ki (cövekfehérjék*).
Nagyon sokféle retrovírus ismert, víruscsaládot (Retroviridae) alkotnak. Sokuk az emberi szervezetre ártalmatlan, mások súlyos betegségeket okoznak: →AIDS (acquired immunodeficiency syndrome), →ATL (adult T-cell leukemia). Ezek – ellentétben a DNS-vírusok átmeneti fertőzéseivel – jóformán megmaradnak a fertőzöttek egész életében, mivel a keletkezett vírus-DNS a sejt-DNS-nek a részévé vált. Kezelésük is életen át tartó.
A csírasejtek retrovírusfertőzéseiből évmilliók során kisebb bázissorok beépültek az emberi DNS-be; annak részévé váltak, és tovább adódtak az utódokba. Ezeket a DNS-részeket retrovíruselemeknek (retroviral elements) nevezzük. (→retrovíruselemek)
(emberi) belretrovírus* (human) endogenous retrovirus, HERV (sokszor csak ERV-nek mondják) öröklött provírus: a retrovírus RNS-éről képződött olyan DNS, amelyik évmilliókkal korábban épült be az emberi DNS-be, és továbbadódik a csírasejtekkel. (→belretrovírusok)