RING (really interesting new gene), RING finger domain/protein RING gomoly az RNS hurkaihoz kötődő gomoly. (→fehérjegomoly)
Találatok címszavakban (1 szócikk):
RING ( really interesting new gene) finger domain RING gomoly (→fehérjegomoly)
Részleges egyezések (15 szócikk):
BARD1 (BRCA1-associated RING domain protein 1) ~780 aminosavas fehérje, az N-végén cinkujj mintázatú gomoly (RING [really interesting new gene] finger domain), a C-végen pedig két BRCT-gomoly van, ezek nagyon hasonlók a BRCA1 fehérjében található gomolyokhoz. Jellegzetes még a három ankirin mintázat. A BARD1 a BRCA1-gyel társul, azt rögzíti, a BRCA1 így fejti ki a hatását. A BARD1–BRCA1 társulást akadályozó másulás következtében a BRCA1 nem tevékeny.
bázispárosodás* base-pairing, Watson–Crick base pairing a nukleinsavak bázisainak kapcsolódása gyenge kötéssel; általában hidrogénhidakkal. A kapcsolódás fajlagos: a DNS-ben az adenozin a timinnel (A–T), a guanin a citozinnal (G–C) párosodik. az RNS-ben az adenozin az uracilhoz kötődik (A–U). Tehát csak az egymáshoz illő bázisok kapcsolódhatnak (illeszkedő párosodás*). A bázispárosodás nemcsak a megfelelőséget biztosítja, hanem rögzíti is a molekulát. A bázisok egyenlő távolságra vannak egymástól, közöttük hézag van; ezt hidalja át a hidrogénkötés. (→DNS)
párhiba base-base mismatch nem illeszkedő bázisok párosodása. (→párhiba)
párosodáshiba egy-két nukleotid vesztése vagy beépülése bázissorba. (→párosodáshiba)
circulating tumor DNA (ctDNA) →keringő daganat-DNS
ctDNA (circulating tumor DNA) →keringő daganat-DNS
DNS-szerkesztés
genome editing, genome engineering,
gene editing (genomszerkesztés)
a DNS genetikai megváltoztatása: beékelünk, kilökünk, módosítunk vagy cserélünk
egy bázist vagy bázissort az élő szervezet DNS-ébe. A DNS-szerkesztés,
ellentétben a véletlenszerű korai genetikai változtatásokkal célzott, sajátos
helyen történik, például célmolekula előállítására. Egyre inkább a kezelések
megerősítésére szolgálnak, annyira, hogy elterjedt az emberi génszerkesztési
kezelés fogalma is. Feltétlen ez a jövő, a rák kezelésében mindenképp. Jó példa
a használatára a sarlósejtes vérszegénység és a Duchenne-kór; ezeknek a
betegségeknek a génhibái genetikai szerkesztéssel gyógyítjuk, de genetikai
szerkesztést alkalmazunk a CART-T-sejtes kezelésekben is.
INO80 (inositol-requiring mutant 80) chromatin-remodeling ATPase. Helikáz (ATPáz) összetevője a kromatint készíti elő a fehérjék kapcsolódására: a magtestecseket távolítja egymástól. Meghatározó a DNS-kettőződés és a DNS-javítás folyamataiban; a rokonmásjavításban állványfehérje.
keringő daganat-DNS (ctDNS) circulating tumor DNA, ctDNA a daganatból vagy a keringő daganatsejtekből származó piciny (~120–180 bázispárnyi [1 magtestecs és az összekötő szál]) DNS-darab a vérplazmában vagy más testnedvben. Származhat az elhaló vagy sejtvégzeti daganatsejtekből, de elválaszthatja az élő daganatsejt is. Nem tévesztendő össze a szérumban szabadon keringő DNS-sel (cell-free DNA, cfDNA). Miután a daganatból származik, tükrözi a daganatsejt DNS-ét, ezért használják génhiba kimutatására is, pl. szokványos a végbél-vastagbél daganatokban a RAS gén vizsgálatára. Alkalmazható továbbá a kezelésre adott válasz megítélésére, valamint a rejtett maradékbetegség (a kiújulás) jelzésére, pl. a kezelést 2–4 héttel követően jelen lévő ctDNS maradékbetegségre utal; ebből újul ki a daganat. A kiújuló daganatból származó ctDNS szerkezete általában más (más génhiba), mint az elsődleges daganatból származóké, utalva a kiújuló daganat fejlődésére. Lévén, hogy a kiújuló daganatból származik, molekulakezelési célpont is lehet, pl. a PD1-gátlók akadályozhatják a kiújuló daganat fejlődését. A ctDNS-t szokásosan a vérből, ritkábban más testnedvből nyerjük ki (liquid biopsy), azonosításához sajátos genetikai vizsgálatokat alkalmaznak. A vérből vett mintának két nagy hátránya van: 1. kevés a ctDNS, 2. keveredik a nem daganatsejtes eredetű keringő DNS-sel (cfDNS, cell-free DNA).
korai TP53-hibás burjánzatok* benign-appearing early serous proliferations(ESPs) with TP53 mutations hámbeli sejtek, amelyekben nem alakulnak ki a hámbeli rákosodás sejtjellemzői, vagy szinte észrevehetetlen mértékben, de hordozzák a TP53 hibát. A hámszövet szerkezetét nem változtatják meg; a génhiba immunfestési módszerével mutathatók ki.
Jelentőségüket az adja, hogy leválhatnak és szóródhatnak a petefészekbe és/vagy a hasüregbe. Ahol megtapadnak, rák keletkezhet belőlük.
körforgás, keringés orbital angular momentum a test körpályán való mozgása; a test egy másik test (középtömeg*) körül mozog, például a Föld keringése a Nap körül. Olyan egyenes irányú mozgás, amely minden pillanatban változtatja az irányát, vagyis a középtömeggel szöget bezáró kerületi mozgás. Ugyanolyan erő hozza létre, mint az egyenes irányú mozgást. A keringő test azért nem halad egyenes pályán, mert egy másik test nem engedi, hogy eltávolodjék, például a tömegvonzás miatt. Ha megszűnik a középtömeghez rögzítő erő, a test a körpályára érintőleges egyenes mentén elrepül.
▪ A keringő test pályája kör. A középtömegtől a keringő testhez húzott sugár a vezérsugár; a test elmozdulásával a vezérsugár is elfordul. Azt a körívszakaszt, amelyet a test megtesz, ívnek nevezzük. Jele: i.
▪ A keringési idő egy kör megtételéhez szükséges idő. Jele: T. Mértékegységes: s (másodperc).
▪ A fordulatszám a keringő test által időegység alatt megtett körök száma. Jele: n. Mértékegysége. 1/s.
Egyenletes a körforgás, ha a test egységnyi idő alatt mindig egyforma nagyságú körívet tesz meg.
lépvérkeringés A lépet a lépütér (splenic/lienal artery) látja el, amely a hasi értörzs (truncus coeliacus) egyik ága. Két–három ágra oszolva a lépkapun hatol a lépbe, és oszlik kisebb ágakra a gerendák szerint (gerendai üterek, trabecular arteries), majd, kilépve a gerendákból, középi üterecskékre (central arterioles) oszlik, amelyekből a tüszői üterecskék (follicular artelioles) erednek. A üterecskék végágai a szegélysávon túl, ecsetszerűen szétágazva nyílnak a vörös lépbélbe; a vér tehát kifolyik a hajszálérszerű végágakból (nyitott keringés).
Az ecsetszerű hajszálerek fala megvastagodott, ami az érhámsejtekhez tapadó nagyfalósejtek sokaságából adódik; a sejtek szinte behüvelyezik az erecskéket (hüvelyes hajszálerek, sheated capillaries, Schweiger–Seidel-hüvely). Ezeknek az erecskéknek a hosszmetszete orsó vagy tojás alakú, ezért orsóereknek vagy ellipszoidoknak nevezik őket. A lépkötegekben a vér a sejtek között áramlik, így a sejtek közvetlenül érintkeznek a vérrel. Innen a vérsejtek és a vérplazma folyamatosan összeszedődik a lépöblökbe a lépöblök nyílásain keresztül.
A lépöblök tekinthetők visszhajszálereknek, amelyekből a vér egyre tágabb visszerekben gyűlik össze. Ezek a visszerek a lépgerendákban haladnak a lépkapu felé, és a lépen kívül egyesülnek; így képződik a lépvisszér (splenic vene), amely a kapukeringésbe ömlik.
Állatokban egyes hajszálerek közvetlenül a lépöblökbe nyílnak (zárt keringés). Az emberi lépben ilyen összeköttetéseket nem mutattak ki. A lép a nyirokkeringéssel nincsen kapcsolatban, benne csak a fehér lépbélből összeszedődő, elvezető nyirokerek vannak.
piRNS (Piwi fehérje-RNS) piwi-interacting RNA, piRNA egyszálú, 23–30 nukleotid hosszú RNS; a kis közbeavatkozó RNS-ek közül a legnagyobb; a piRNS géncsoportosulások (piRNA clusters) terméke. Ezeket a géneket az RNS-polimeráz-II írja át; hosszú folytonos egyszálú RNS (ssRNA; single strand) keletkezik. Ez kerül a sejtplazmába, ahol RNáz hasítja piRNS-sé, vagy a már meglévő piRNS-t tartalmazó RISC (piRISC) köti bázispárosodással, és hasítja annak megfelelően.
Mindkét folyamatban 5’-monofoszfát közbenső piRNS (5’ monophosphate piRNA intermediate) képződik, amelyik hosszabb az érett piRNS-nél, és kapcsolódik PIWI fehérjével; ez alakítja érett piRNS-sé (→RISC).
Három alcsaládja ismert: a rasiRNS, a crasiRNS és a tel-sRNS – tevékenységük csak részben ismert.
rasiRNS (repeat-associated small interfering RNAs, rasiRNA) a férfi ivarsejtekben szabályozza az ugrálatok (transposons) és a visszugrálatok (retrotransposons) sokaságát.
crasiRNS (centromere repeat associated small interfering RNA) középrész-RNS* a kromoszómák középrészének ismétleteiről íródnak át, tevékenységük alig ismert.
telsRNS (telomere-specific small RNA, tel-sRNA) végrész-RNS* a kromoszómák végeinek, a végrészismétleteknek az átíródásából keletkező kis RNS (végrészátíratok, telomeric transcripts). A tel-sRNS-eket az ébrényi (embryonal) őssejtekben fedezték fel. Szerepük kevéssé ismert, bizonyára a végrészek kromatinjának szervezésében vesznek részt.
Az emberben a piRNS csak piRISC formájában van jelen. A piRNS-ek működésének megismerése további vizsgálatokat igényel. Jellegzetes, hogy a heterokromatikus területeken vannak jelen, feltehetően a heterokromatkus részek alapvető szabályozói. (→RISC)
siRNS (small interfering RNA, siRNA) kis közbeavatkozó RNS* (egyéb nevei: short interfering RNA, silencing RNA) 19–25 nukleotidból álló, kétszálú RNS. Az miRNS-re jellemző szárhurok nincs benne. Két egyszálú RNS párosodik; a szálak a 3’-végen túlnyúlnak két bázissal. Az egyik szál az irányítószál (guide strand, antisense strand) a másik a kísérőszál (passenger strand, sense strand). (→miRNS)
A sejtplazmába került hosszú kétszálú (egyenes vagy szárhurkot tartalmazó) RNS-ből a DICER hozza létre az érett siRNS-t, lehasítva a hosszú kétszálú RNS 3’-végét. Hasonlóan az miRNS-hez, kapcsolódik AGO-fehérjével, kialakítva a RISC-et. A társulás következtében szétválik a siRNS két szála; a kísérőszál lebomlik. Az irányító szál vezeti a RISC-et a kiegészítő bázispárok szerint azonosított mRNS-ekhez. Azokkal tökéletesen párosodik, ezért az siRISC mindig hasítja az mRNS-t.
A sejtplazmába kerülő hosszú kétszálú RNS-ek keletkezhetnek a sejtben (belső siRNS, endo-siRNA), de bekerülhetnek a sejtbe a sejt környezetében lévő hosszú kétszálú RNS-ek is (külső siRNS, exo-siRNA); pl. a vírus mRNS-ből létrejövő kétszálú RNS alakul siRNS-sé.
Hozzávetőleg 200-féle siRNS-t ismerünk. Az siRNS is a RISC tagjaként szabályoz. Az siRNS-t kiterjedten alkalmazzák a ráksejtek elpusztítása: egyszerűem beadható (kivált a belégzési kezelés ígéretes; az érbe adva az enzimek gyorsan bontják), a ráksejtek burjánzását előidéző géneket célzottan gátolja, nincs mellékhatása.
szerkezeti azonmások* constitutional isomers azonos atomösszetételű és molekulatömegű, de az atomok kapcsolódásában (összekapcsolódási rendjében) eltérő molekulák. Másként: azonos összegképletű, de különböző szerkezeti képletű vegyületek. A szerkezeti azonmások fizikai tulajdonságai, pl. forráspontjuk, különbözik. (→azonmás)
lánc azonmások* chain isomers az atomok, szokásosan a szénatom elágazásában és/vagy kötésükben eltérő azonmások. Pl.
sorrendi eltérés:
n-bután:
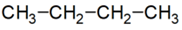
izobután:
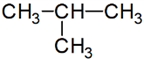
kötési eltérés:
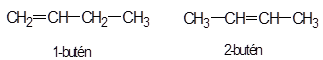
A legegyszerűbb szénhidrogéneknek, mint metán (CH4), etán (CH3–CH3) és propán (CH3–CH2–CH3), nincs azonmása, mert a szén és hidrogének kapcsolódása csak egyféleképpen lehetséges.
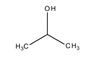
 hatócsoportú
azonmások* functional (group) isomers
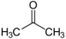
különböző hatócsoportot kapcsoló azonmások. Pl. az acetonban karbonil- (C=O), a
propanolban pedig hidroxilcsoport van.
hatócsoportú
azonmások* functional (group) isomers
különböző hatócsoportot kapcsoló azonmások. Pl. az acetonban karbonil- (C=O), a
propanolban pedig hidroxilcsoport van.
kötésváltó azonmások* tautomers csak a protonok és elektronok elhelyezkedésében más azonmások. A kötésátalakulás voltaképpen protonáthelyeződés* (prototropy): egy proton a molekula egyik helyéről a másikra tevődik át, rendszerint az elektronok eloszlási változásával, a mozgékony hidrogén áttevődésével és kötésváltozással jár. Ezek egyféle kötésben eltérő vegyületek, amelyek könnyen átalakulnak egymásba. Együtt vannak jelen, váltakozó egyensúlyi állapotban, és a vegyi folyamatokban szintén együtt vesznek részt. A kötésváltásnak* (tautomerism) számos formája ismert. Néhány gyakoribb:
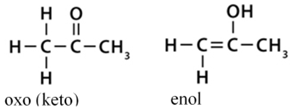 • oxo(keto)–enol azonmásság*. Az α helyzetű szénatom hidrogénjének
áthelyeződése. Az oxovegyületekben egy oxigénatom kettős kötéssel kapcsolódik
egy szénatomhoz. Az enolformában OH-csoport kötődik telítetlen szénvegyülethez.
Az oxovegyületek előfordulhatnak
enolformában is (oxo–enol kötésváltás). Az oxoforma erősebb kötésű, és túlsúlyban van a kettő egyensúlyi
állapotában. (→hatáscsoport, oxovegyületek)
• oxo(keto)–enol azonmásság*. Az α helyzetű szénatom hidrogénjének
áthelyeződése. Az oxovegyületekben egy oxigénatom kettős kötéssel kapcsolódik
egy szénatomhoz. Az enolformában OH-csoport kötődik telítetlen szénvegyülethez.
Az oxovegyületek előfordulhatnak
enolformában is (oxo–enol kötésváltás). Az oxoforma erősebb kötésű, és túlsúlyban van a kettő egyensúlyi
állapotában. (→hatáscsoport, oxovegyületek)
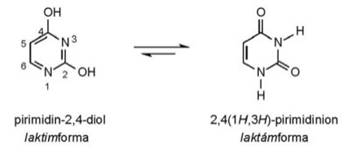 • laktim–laktám azonmások. Az α helyzetű nitrogénatom
hidrogénjének áthelyeződése: –N=C– / –N–C–;
a laktimban a szén kettős kötéssel, a laktámban egyes kötéssel kapcsolódik a
nitrogénhez. Gyűrűs vegyületekben fordul elő, a
gyűrűs savamidokra (laktámokra) jellemző a hidrogénátrendeződés a
gyűrűben lévő amidcsoporton belül. Az uracil
(pirimidin-2,4-diol) egyensúlyi folyamata:
• laktim–laktám azonmások. Az α helyzetű nitrogénatom
hidrogénjének áthelyeződése: –N=C– / –N–C–;
a laktimban a szén kettős kötéssel, a laktámban egyes kötéssel kapcsolódik a
nitrogénhez. Gyűrűs vegyületekben fordul elő, a
gyűrűs savamidokra (laktámokra) jellemző a hidrogénátrendeződés a
gyűrűben lévő amidcsoporton belül. Az uracil
(pirimidin-2,4-diol) egyensúlyi folyamata:
• Egyéb kötésváltó azonmások: enamin–imin: H−N−C=C / N=C−C−H, nitrozo–oxim: H−C−N=O / C=N−O−H, keten–inol: H−C=C=O / C≡C−O−H, foszfit–foszfonát: P(OR)2(OH) ⇌ HP(OR)2(=O), aminosav–ammonium-karboxilát (H2N−CH2−COOH / H3N+−CH2−CO−) stb.
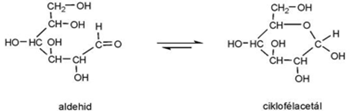 gyűrű–lánc azonmások* ring-chain isomers az azonmások egyike nyíltlánc, a másik gyűrűs. Olyan
aldehid-/ketocsoportot és OH-csoportot tartalmazó vegyületekben, például
szénhidrátokban fordul elő, amelyekben a proton áthelyeződésével a nyílt
vegyületből (láncforma) gyűrűs lesz, és fordítva. Az OH-csoport hidrogénje az
aldehidcsoportra vándorol, kialakítva a gyűrűformát. Az aldehidlánc utolsó
szénatomjához kettős kötéssel kapcsolódó oxigén helyett egyes kötéssel
OH-csoport társul. Az ábrán a glükóz (szőlőcukor) példája látható.
gyűrű–lánc azonmások* ring-chain isomers az azonmások egyike nyíltlánc, a másik gyűrűs. Olyan
aldehid-/ketocsoportot és OH-csoportot tartalmazó vegyületekben, például
szénhidrátokban fordul elő, amelyekben a proton áthelyeződésével a nyílt
vegyületből (láncforma) gyűrűs lesz, és fordítva. Az OH-csoport hidrogénje az
aldehidcsoportra vándorol, kialakítva a gyűrűformát. Az aldehidlánc utolsó
szénatomjához kettős kötéssel kapcsolódó oxigén helyett egyes kötéssel
OH-csoport társul. Az ábrán a glükóz (szőlőcukor) példája látható.
szerves vegyületek organic compounds szénhidrogének; szén- és hidrogénatomokból épülnek fel, de lehetnek bennük más atomok is (oxigén, nitrogén, kén, foszfor stb.), amelyeket idegen atomoknak nevezünk. Az atomok elektronkötéssel kapcsolódnak egymáshoz. A szerves vegyületek egyedülálló tulajdonsága, hogy más szénatomokkal, vagy a szerves vegyületekben előforduló más atomokkal, változatos szerkezeteket (láncok, elágazások, gyűrűk stb.) hoznak létre. Ezek alkotják az élő szervezetet. A szerves vegyületek száma szinte határtalan, több millió ismert, szemben a néhány százezer szervetlen vegyülettel. A természetben előfordul néhány olyan szénatomos vegyület, amelyet nem tartunk szervesnek; ilyenek a karbidok, karbonátok és a cianidok. (→vegykötések)
A szerves vegyületeket háromféle tulajdonság alapján osztályozzuk: aromás és nem aromás, nyíltláncú és gyűrűs, valamint telített és telítetlen szerves vegyületekre.
aromás vegyületek aromatic compounds síkalkatú, egyes és kettős kötésű, hat szénatomos állékony gyűrűs (benzolgyűrű) vegyületek jellegzetes elektronszerkezettel. A kötések mindegyike részt vesz az elektronok mozgásában: 3 pár π-elektron egyenletesen kering minden atom körül (a Hückel-szabály szerint). Az aromás vegyületek telített vegyületként viselkednek; helyettesítő vegyülékenységűek, tulajdonságaik merőben mások, mint az azonos összetételű nyílt láncúaké. (→Hückel-szabály)
gyűrűs (zárt láncú) vegyületek closed (cyclic, ring) compounds szénatomokat tartalmazó gyűrűt képező vegyület, az
első és az utolsó szénatom kapcsolódik egymáshoz. Lehetnek egygyűrűs
(monocyclic compound), kétgyűrűs (bicyclic compound), háromgyűrűs (tricyclic
compound), néhánygyűrűs (oligocyclic compound), sokgyűrűs (polycyclic compound)
összetételűek. A gyűrűk közös atomja szerint lehetnek: 1. elkülönült többgyűrűs
– nincs közös atom; 2. egy közös atomú; 3. két közös atomú és 4. áthidalt
gyűrűs – több közös atomú – vegyületek. Megkülönböztetünk csak szénatomos
gyűrűs vegyületeket (homocyclic [isocyclic] compounds] – a gyűrű valamennyi
tagja szénatom (homociklusos vegyület) – és vegyes gyűrűs vegyületeket
(heterocyclic compounds) – a gyűrűben a szénatomok mellett más atom is van (heterociklusos
vegyület). Mindegyik lehet aromás és nem aromás.
nem aromás vegyületek (alifás vegyületek) minden olyan szerves
vegyület, amelyik nem aromás tulajdonságú. Lehet nyílt láncú és gyűrűs
(alicyclic compounds), valamint telített és telítetlen.
nyílt láncú vegyületek open-chain (acyclic, alophatic) compounds elektronkötésekkel kapcsolódó szénatomok láncából álló szerves vegyületek. A lánc lehet egyenes (linear) és elágazó (branching). Egyszeres (C–C), kettős (C═C) és hármas (C≡C) kötésű szénatomokat (C) tartalmazhatnak.
telítetlen vegyületek unsaturated compounds legalább egy kettős vagy hármas kötéssel kapcsolódó szénatomokat tartalmazó vegyületek. Vegyülékenyek: a kettes/hármas kötés könnyen cserélődik egyesre. Főleg egyesülnek és sokszorozódnak.
telitett vegyület saturated compound a szénatomok mindegyike egyes kötéssel kapcsolódik, nincs szabad kötőhely. Kevéssé vegyülékeny molekulák; főleg helyettesítési folyamatokban vesznek részt. Lehetnek nyílt- és zártláncúak.
zárt láncú (gyűrűs) vegyületek closed (cyclic, ring) compound szénatomos gyűrűt tartalmazó vegyületek. (→szerves vegyületek)