bázishármas codon a DNS-nek vagy RNS-nek egyfajta aminosavat meghatározó
hármas bázissora (nukleotidsora). 64 különböző bázishármas van (43 =
64 változat); ebből 61 felel meg valamely aminosavnak, három pedig az átíródás
befejezésének jelzésére szolgáló záró bázishármas. Ezek alkotják a genetikai
kódot, amely az élővilág minden tagjában egyforma. Mivel jóval több bázishármas
van (bázishármastöbblet), mint aminosav, egyféle aminosavat különböző
bázishármasok is kódolhatnak. Kivétel a metionin és a triptofán; ezeket csak
egyféle bázishármas képez: a metionint az AUG (adenin, uracil, guanin –
megfelelője a DNS-en az ATG [adenin, timin guanin]), a triptofánt az UGG
(uracil, guanin, guanin – megfelelője a DNS-en az TGG [timin guanin, guanin]).
A bázishármastöbblet a fajfejlődés során, évmilliók
alatt alakult ki, valószínűleg összefüggésben az aminosavak fehérjékbeli
előfordulásának gyakoriságával: minél gyakoribb valamely aminosav a
fehérjeösszesben, annál több bázishármas képezi, pl. a leucint, amely a
leggyakoribb aminosav a fehérjékben, hatféle bázishármas (TTG, TTA, CTT, CTC,
CTG and CTA) kódolja. A bázishármastöbblet jelenségét a nemzetközi irodalom degeneracy
of genetic code elnevezéssel illeti.
indító bázishármas start codon az
mRNS első hármasa, amely mindig metionint kódol; legtöbbször AUG. A metionin a
más aminosavval kezdődő fehérjékről később leszakad.
kezdő bázishármas az átíródás kezdetének az első három bázisa
(nukleotidja). A polimeráz ettől olvassa a mintafelületet. Szokványosan az 5’
UTR-t követi. (→génszerkezet, olvasási keret)
záró bázishármas stop codon a DNS-nek olyan hármasa, amely
megállítja az elő-mRNS keletkezését, illetve az mRNS-nek az a bázishármasa,
amely megállítja az átfordítódást; vagyis a fehérjeképződést megállító hármas
bázis, pontosabban nukleotid. Mivel nem kódol aminosavat, nonsense codonnak
is nevezik. Háromféle ismert: UAA, UAG és UGA.
bázispár basepair a
DNS-ben, RNS-ben hidrogénkötésekkel kapcsolt két bázis. A DNS-ben alapvetően az
egymással szembeni kiegészítő két bázis, az adenin és
a timin, illetve a guanin és citozin alkot párt, ezek kötik össze a DNS két
szálát. Szokásosan bázispáron ezeket értjük, nevezik őket Watson–Crick-féle
bázispároknak is.
Bázispár azonban létrejöhet másként, pl. ha
a purinbázis elfordul, az 5-ös gyűrűje kapcsolódik pirimidinhez, nem pedig a
hatos gyűrű, avagy kép purinbázis között. Ezeket Hoogsteen-féle bázispároknak
(Hoogsteen-bázisporoknak) nevezzük; az őket összekötő hidrogénkötések a
Hoogsteen-féle hidrogénhidak. (→hidrogénkötés)
A
kétszálú DNS/RNS hosszát bázisokkal (b), bázispárokkal (bp) fejezzük ki
számokkal vagy kilo, mega-, giga- mértékegységgel: 1000 bázis / 1000 bázispár =
1 kilobázis (kb) / 1 kilobázispár (kbp). 1 000 000 bázis / 1 000 000
bázispár = 1 megabázis (Mb) / 1 megabázispár (Mbp).
1 000 000 000 bázis / 1 000 000 000 bázispár = 1
gigabázis (Gb) / 1 gigabázispár (Gbp). Az egyszálú DNS/RNS-nél a nukleotidok
számát adjuk meg. Jele: nt (knt – kilonukleotid, Mnt – meganukleotid, Gnt –
giganukleotid).
bázispárcsere* single
nucleotide variation, SNV egyetlen bázispár cserélődése másik illeszkedő
bázispárra a DNS bázissorában. Lehet bázishasonló*, amikor purinbázis
cserélődik purinbázisra, illetőleg pirimidinbázis pirimidinbázisa; és lehet
báziselütő*: purinbázis–pirimidinbázis csere. A cserélődött bázis kiegészül az
illeszkedő párjával. A bázispárcsere a DNS bármely részén előfordul; leginkább
a gének között, de a gének kódoló (1 százalék) és nem kódoló részében is.
Különösen gyakori az ismétletekben. A bázispárcseréből alakult bázissor a bázispárcserés
változat (single nucleotide variant, SNV).
A
kódoló szakaszban lévő bázispárcseréből keletkező fehérje (aminosavsor) formája
szerint háromféle ismert:
azonaminosavas párcsere* synonymous
mutation (nevezik single amino acid samesense change-nek is) olyan
báziscserélődés, amely nem változtatja meg a fehérje aminosavait. Keletkezhet
ugyanis olyan bázishármas, amelyik ugyanazt az aminosavat kódolja, például
GTT>GTC változás; mindkettő valint kódol. Az emberi DNS-ben ugyanis 64
bázishármas kódol 20 aminosavat, azaz egy-egy aminosavat több bázishármas is
képezhet. Ezt a jelenséget a nemzetközi irodalom degeneracy of genetic code-nak
nevezi, magyarul génkódbőségnek* mondhatjuk. A polipeptid képződése vagy
alakulása azonban módosulhat, például változhat az átíródás, átfordítódás
üteme, a közteskivágás stb., ezért az egyedségben valamilyen módon
megnyilvánul.
néma azonaminosavas párcsere* silent mutation
(néma másulás, néma mutáció) olyan aminosav-változást nem okozó egyetlen
bázispár-cserélődés, amely az egyedségben jóformán nem nyilvánul meg. A polipeptidlánc
képződése és további alakulása változatlan.
 egy aminosavas párcsere* missense mutation, non-synonymous mutation (más néven single amino acid missense change,
missense mutation) egyetlen aminosavcserét okozó
bázispár-cserélődés: a polipeptidlánc egyik aminosavának helyére másik épül be.
Szabálytalanul (másként) működő vagy nem működő fehérje keletkezik. Az ábrán a
prolint, a glicint és a tirozint kódoló bázishármas látható. A TA bázispár
cserélődik GC-re, aminek következtében nem tirozint, hanem szerint kódoló
bázishármas alakul ki az mRNS-en. A keletkező fehérjében egy tirozin helyett
szerin lesz. A polipeptid további képződése változatlanul folytatódik.
Összetételében más fehérje keletkezik, de az aminosavsor és a fehérje hossza
sem változik. A létrejövő fehérje működése megváltozhat, változhat a szerkezete
is. Ez elsősorban attól függ, hogy milyen aminosav cserélődik: ha lényegesen
más tulajdonságú épül be, mint az eredeti, nagyobb zavar keletkezhet a fehérje
működésében.
egy aminosavas párcsere* missense mutation, non-synonymous mutation (más néven single amino acid missense change,
missense mutation) egyetlen aminosavcserét okozó
bázispár-cserélődés: a polipeptidlánc egyik aminosavának helyére másik épül be.
Szabálytalanul (másként) működő vagy nem működő fehérje keletkezik. Az ábrán a
prolint, a glicint és a tirozint kódoló bázishármas látható. A TA bázispár
cserélődik GC-re, aminek következtében nem tirozint, hanem szerint kódoló
bázishármas alakul ki az mRNS-en. A keletkező fehérjében egy tirozin helyett
szerin lesz. A polipeptid további képződése változatlanul folytatódik.
Összetételében más fehérje keletkezik, de az aminosavsor és a fehérje hossza
sem változik. A létrejövő fehérje működése megváltozhat, változhat a szerkezete
is. Ez elsősorban attól függ, hogy milyen aminosav cserélődik: ha lényegesen
más tulajdonságú épül be, mint az eredeti, nagyobb zavar keletkezhet a fehérje
működésében.
 záróhármashiba* nonsense mutation záró bázishármas* (stop
codon) kialakulásához vezető egyetlen bázispár cserélődése, amelynek
következtében csonkfehérje* (truncated protein) keletkezik. A fehérje egy része
hiányzik, és általában nem működik. Szerkezete attól függ, hogy a báziscsere
következtében melyik bázishármas alakul záróvá. Az ábrán a prolint, a glicint
és a tirozint kódoló bázishármas látható. A GC bázispár csrélődik CG-re, aminek
következtében nem tirozint kódoló hármas, hanem záróhármas (UAG) keletkezik az
mRNS-en, és az átfordítódás idő előtt befejeződik, hiányos fehérje jön létre.
záróhármashiba* nonsense mutation záró bázishármas* (stop
codon) kialakulásához vezető egyetlen bázispár cserélődése, amelynek
következtében csonkfehérje* (truncated protein) keletkezik. A fehérje egy része
hiányzik, és általában nem működik. Szerkezete attól függ, hogy a báziscsere
következtében melyik bázishármas alakul záróvá. Az ábrán a prolint, a glicint
és a tirozint kódoló bázishármas látható. A GC bázispár csrélődik CG-re, aminek
következtében nem tirozint kódoló hármas, hanem záróhármas (UAG) keletkezik az
mRNS-en, és az átfordítódás idő előtt befejeződik, hiányos fehérje jön létre.
A nem kódoló DNS-szakaszon lévő
bázispácsere szintén befolyásolhatja a fehérjeképződést: hatással lehet a
mRNS-re, a nem kódoló RNS-re, az átírásfehérje kapcsolódására, az átfordításra
és a közteskivágásra is.
Az egy aminosavas párcserét és a záróhármashibát közös néven másaminosavas párcserének (nonsynonymous SNV) nevezzük.
bázispárcserés változat*
single nucleotide variant, SNV a DNS-nek olyan bázissora, amelyben
egyetlen bázispár másik illeszkedő bázispárra cserélődött. Miként a
bázispárcsere, a bázispárcserés változat is lehet bázishasonló és báziselütő,
előfordulhat a DNS bármely részén, valamint lehetnek azonaminosavas, egy
aminosavas és záróhármashibás változatok. A kódoló és szabályozó bázissorokban
lévő változatok befolyásolhatják a génkifejeződést, megváltoztathatják a
fehérjék közötti kölcsönhatásokat, módosíthatják a fehérjéket, hatással
lehetnek a sejtműködésre, megnyilvánulhatnak az egyedségben; például betegséget
okoznak. A DNS más részeiben előfordulók rendszerint közömbös változatok, ám
ezek között is lehetnek az egyedségre hatók, például betegségre hajlamosítók,
avagy éppen a betegség kialakulását gátlók.
A DNS-ben ~10 millió bázispárcserés változat van;
100–300 bázisonként fordul elő. Legtöbbször (háromból kettőben) a citozin
cserélődik timinre.
A bázispárcserés változatok meglehetősen állandók,
megmaradnak nemzedékről nemzedékre, ezért népességi tanulmányokban jól
követhetők, nagyon alkalmasak genetikai összefüggések tisztázására, például a
genetikai betegségekkel összefüggő térképezésére, betegségre hajlamosító
báziseltérések, genetikai jelleg meghatározására stb.
bázispárosodás* base-pairing,
Watson–Crick base pairing a nukleinsavak
bázisainak kapcsolódása gyenge kötéssel; általában hidrogénhidakkal. A
kapcsolódás fajlagos: a DNS-ben az adenozin a timinnel (A–T), a guanin a
citozinnal (G–C) párosodik. az RNS-ben az adenozin az uracilhoz kötődik (A–U). Tehát
csak az egymáshoz illő bázisok kapcsolódhatnak (illeszkedő párosodás*). A
bázispárosodás nemcsak a megfelelőséget biztosítja, hanem rögzíti is a
molekulát. A bázisok egyenlő távolságra vannak egymástól, közöttük hézag van;
ezt hidalja át a hidrogénkötés. (→DNS)
párhiba base-base
mismatch nem illeszkedő bázisok párosodása.
(→párhiba)
párosodáshiba egy-két nukleotid vesztése
vagy beépülése bázissorba. (→párosodáshiba)
bázispásztázás* sequencing a bázisok egyenkénti
vizsgálata a DNS-ben, RNS-ben.
▪ Sanger-bázispásztázás* (Sanger sequencing).
A teljes bázissor átnézésének legrégebbi módszere. Előnye, hogy a vizsgálat
alapjául szolgáló láncmegállítás a bázissorrend igen pontos meghatározását
teszi lehetővé. Hátránya, hogy egyszerre legfeljebb 600 bázispárnyi szakaszt
képes vizsgálni, így a teljes szakasz elemzése nagyon sok időt vesz igénybe. A
módszer érzékenysége ~10−15%, tehát a kimutatni kívánt eltérésnek a mintában
legalább ilyen arányban jelen kell lennie. Más módszerrel kimutatott génhibák
megerősítésére is használják.
▪ Újnemzedékes bázispásztázás* (next
generation sequencing, NGS). Az új, gyorsabb
bázispásztázó módszerek kifejlesztése a XXI. század elejétől kezdődött, és
módszertanilag igen szerteágazó. Az NGS gyűjtőfogalom: különböző cégek többféle
vegyi módszeren alapuló eljárásait jelenti. Az NGS lehetővé teszi nagyméretű
DNS-szakaszok gyors átnézését, érzékenysége pedig elérheti akár az 1−2%-ot is.
A BRCA1 és 2 gének vizsgálatához már számos előre gyártott készlet létezik. A
módszer hátránya, hogy nagyon nagy mennyiségű adat keletkezik, melynek
értelmezése komoly szakértelmet kíván. Az NGS-eredmények értelmezésének
megkönnyítésére különböző alkalmazásokat fejlesztenek ki, mint például a BRCA-elemző
(analyzer). Ez olyan önműködő munkafolyamat, mely a BRCA1 és 2 gének NGS-adatainak
elemzését végzi. Az alkalmazás ingyenesen hozzáférhető.
▪ Az emberi DNS teljes
bázispásztázása (whole genome sequencing, WGS) olyan nagy felbontású
módszer, amely képes lesz felváltani a kromoszómák szokványos vizsgálati módjait.
A báziseltéréseken kívül a
módszer alkalmas a kromoszómák szerkezeti elváltozásainak (másolatszám-változás,
kiegyensúlyozott átrendeződés) kimutatására is.
bázissor sequence a DNS-ben, RNS-ben a bázisok,
nukleotidok egymásutánja. Ez határozza meg a működésüket. Változás a
bázissorban változást hozhat, általában hoz is, változást a DNS vagy RNS tevékenységében.
■ Törzs bázissor (consensus vagy canonical sequence) a bázissor
valamelyik részén leginkább előforduló bázissor; az a változata, amelyhez
hasonlítjuk a többit. Szokásosan ezt tartjuk az ép bázissornak. Számításokkal
határozzák meg.
bázissoróra* epigenetic
clock néhány száz sajátos CpG-hely metilezettségi változásának a mértéke.
Segítségével pontosan meghatározható a biológiai életkor, összevethető a valós,
években mért életkorral, és jelzője lehet betegségeknek is.
bázistöbblet*, bázisvesztés* insertion, deletion. Általános
értelmezésben a bázistöbblet* egy vagy több bázis (pontosabban nukleotid)
beékelődése, a bázisvesztés* egy vagy több bázis (nukleotid) törlődése.
Előfordulhat a DNS bármely részén; gyakori az ismétletekben.
Ha beékelődik vagy törlődik egy vagy több bázis (nukleotid), a DNS-szál hurokszerűen
kiboltosodik; ezt nevezzük beékelődési/törlődési huroknak* (insertion–deletion loop,
IDL).
A bázistöbbletnek/bázisvesztésnek három formája ismert: a kereteltolódás, az INDEL
(bázistöbblet/bázisvesztés*) és a szakaszkettőződés.
 ▪ Kereteltolódás*
(frameshift mutation) az
olvasáskeretnek egyetlen bázis beékelődése vagy kiesése miatti olyan másulása,
amelyben megváltozik az aminosavat kódoló bázishármasok sorrendje. Teljesen más
fehérje keletkezik, amely nyilvánvalóan nem alkalmas az eredeti fehérje
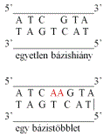
feladatának ellátására. Az ábra felső részén öt CAT bázishármas látható; öt
hisztidin keletkezik, és képződik tovább a polipeptid. Az alsó bázissorban a
második hármas után beékelődik egy adenin (A), és teljesen átalakul az
olvasáskeret: megváltoznak a bázishármasok. A harmadik hármas treonint, a
negyedik és ötödik szerint képez és így tovább. A képződő fehérje szerkezete
tehát egészen más lesz.
▪ Kereteltolódás*
(frameshift mutation) az
olvasáskeretnek egyetlen bázis beékelődése vagy kiesése miatti olyan másulása,
amelyben megváltozik az aminosavat kódoló bázishármasok sorrendje. Teljesen más
fehérje keletkezik, amely nyilvánvalóan nem alkalmas az eredeti fehérje
feladatának ellátására. Az ábra felső részén öt CAT bázishármas látható; öt
hisztidin keletkezik, és képződik tovább a polipeptid. Az alsó bázissorban a
második hármas után beékelődik egy adenin (A), és teljesen átalakul az
olvasáskeret: megváltoznak a bázishármasok. A harmadik hármas treonint, a
negyedik és ötödik szerint képez és így tovább. A képződő fehérje szerkezete
tehát egészen más lesz.
 ▪ INDEL (
in = insertion,
del
= deletion) magyarul bázistöbblet/bázisvesztés*, de egységesség kedvéért
az INDEL szóösszevonást alkalmazzuk. Megegyezés szerint az INDEL legtöbb 1 kb
hosszúságú (1–1000 bázispár) bázistöbblet/bázisvesztés. Helytelen rövid/hosszú
többletről/vesztésről (small/large indel) beszélni, mert a fogalom jelentése
szerzők szerint nagyon is eltérő. Ha szükséges, pontosítani kell a
törlődött/beékelődött bázisok számát. Az egyetlen bázispár
beékelődése/törlődése pontmásulás. Egyik formája a kereteltolódás. A bázisok
százait tartalmazó beékelődéseket/törlődéseket, például kromoszómaáthelyeződés/kromoszómamegfordulás,
szintén INDEL-nek nevezzük, mert nem külön másulási forma.
▪ INDEL (
in = insertion,
del
= deletion) magyarul bázistöbblet/bázisvesztés*, de egységesség kedvéért
az INDEL szóösszevonást alkalmazzuk. Megegyezés szerint az INDEL legtöbb 1 kb
hosszúságú (1–1000 bázispár) bázistöbblet/bázisvesztés. Helytelen rövid/hosszú
többletről/vesztésről (small/large indel) beszélni, mert a fogalom jelentése
szerzők szerint nagyon is eltérő. Ha szükséges, pontosítani kell a
törlődött/beékelődött bázisok számát. Az egyetlen bázispár
beékelődése/törlődése pontmásulás. Egyik formája a kereteltolódás. A bázisok
százait tartalmazó beékelődéseket/törlődéseket, például kromoszómaáthelyeződés/kromoszómamegfordulás,
szintén INDEL-nek nevezzük, mert nem külön másulási forma.
Bázistöbblet/bázisvesztés előfordul az olvasáskeretben
az olvasáskeret eltolódása nélkül is. Ezt a nemzetközi irodalom in frame
insertion/deletion nevezettel illeti. Magyarul: keretmegtartó
többlet/vesztés* Három vagy hárommal osztható bázisok többletével vagy
törlődésével jön létre. Például: három bázis kiesése egy aminosav vesztésével
jár, a további kódoló hármasok azonban változatlanul megmaradnak, nem tolódik
el az olvasáskeret.
INDEL többféleképpen
keletkezhet: lehet DNS-károsodás (sugárhatás stb.) következménye, de
kialakulhat a DNS-polimeráz hibájából (szálcsúszás), avagy téves, nem megfelelő
DNS-javítás miatt.
▪ Szakaszkettőződés* általában 1–200 kb nagyságú DNS-rész egynél többszöri előfordulása a
DNS-ben; a DNS bármely elemét tartalmazhatja. Az eredeti és az ismétlődő
szakasz szerkezete >90 százalékban azonos. A szakaszkettőződés az emberi DNS
~5 százalékában fordul elő. A kettőződött szakaszok hajlamosak újabb
ismétlődésekre, szerkezeti átalakulásokat eredményezve, amelyek a kromoszómákon
sokszor jól felismerhetők, sőt jelentősen megnyújthatják a kromoszómát. Ha az
ismétlődő részben gén is van, két, több egyazon génje is lehet az egyébként
egészséges embernek. Ám ha a gén tevékenysége fokozódik (gene dosage imbalance)
a gének többlete miatt (mennyiségérzékeny gén), betegség keletkezhet.
~60 000-féle szakaszkettőződés ismert. Nagyságrendje miatt a
szakaszkettőződés a DNS-nek méretesebb eltérése, mint az egynukleotidos
sokalakúság összessége.
A szakaszismétlődés a génkifejlődés, új gének
keletkezésének alapja, például a törzsfejlődésben vagy a környezeti
alkalmazkodás folyamataiban. A kettőződés következtében rendelkezésre áll
bázissor, amelyből új gén keletkezhet, például úgy, hogy szabályozó részek is
másolódhatnak, és alakíthatják a bázissort, vagy a nem válzati kereszteződések
(non-allelic homologous recombination) folyamatában. A bázissor átalakulása
szakaszos, az új gén több ismétlődésben jön létre. Nemrégiben felismert
szakaszismétlődésből keletkezett új gén az ARHGAP11B és a SRGAP2C; ezeknek az
agykéreg és a gerincvelői idegek fejlődésében van szerepük.
A kettőződött, többszöröződött szakaszok egymás után
ismétlődnek, ezért az ismétletek közé is sorolják őket (low copy repeats). Az
ismétletektől azonban lényegileg különböznek, szoros értelemben nem tartoznak
közéjük.
A bázistöbblet/bázisvesztés vizsgálata nemcsak a laboratóriumi
módszerekben jelentős, hanem az orvosi gyakorlatban is: örökletes vagy szerzett
betegség megkülönböztetésében, avagy egyes betegségek kórismézésében, például
háromnukleotidos ismétlettöbblet bántalom (trinucleotide repeat expansion
diseases), valamint kezelésben, kórjóslatban, például Lynch-kór.
báziszsúfolódás* base-stacking
az a természetes folyamat, melyben az ugyanazon a DNS szálon helyet foglaló
szomszédos nukleotidok bázisai lap szerint egymás fölé rendeződnek. Az így
egymásra halmozott, angolul stacked, bázisok között jellemzően gyenge
kölcsönhatások által közvetített kapcsolatok alakulnak ki, amelyek rögzítik az
egyes szálú nukleotidsor térszerkezetét.
A hidrogénhidakkal a bázisok úgy párosodnak, hogy
köztük hézag van. A bázisok felszínén töltéses kötések vannak, ezért a bázis
víztaszító. Ebből adódik, hogy a vizes közeg taszítja a bázist, a bázispárokat
távolítja, csökkentve a molekula állékonyságát. A bázisok zsúfolódásával
csökken a vizes közeggel érintkező felszín, a taszító hatás jóval kevésbé
érvényesül. sokkal kevesebb energia kell a bázisok összetartásához: a molekula
szerkezete állandósul.
A báziszsúfolódás a nukleinsavak, kivált a kétszálú
nukleinsavak legerősebb összetartó ereje. Ennek különösen vizes közegben
(sejtplazma) van jelentősége. A bázistömörödést a bázisok módosulása (pl.
metilezés) alakítja: a módosulás következtében elfordul a gerinc, megváltozik a
bázisok viszonya egymáshoz, közelebb kerülőket gyenge kötések, főleg a van der
Waals-erő összébb húzza.
gyenge bázis vizes oldatban csekély mértékben szétváló bázis.
(→bázis)
 purinbázis purine base purinszármazék, nitrogént
tartalmazó kétgyűrűs vegyület, a nukleinsavak építőeleme. A DNS-ben és az
RNS-ben is megtalálható az adenin (6-amino-purin) és a guanin
(2-amino-6-hidroxi-purin). A nukleinsavakban a pirimidinbázisokkal társulnak
hidrogénkötésekkel: az adenin a citozinnal, a guanin a DNS-ben a timinnel, az
RNS-ben az uracillal.
purinbázis purine base purinszármazék, nitrogént
tartalmazó kétgyűrűs vegyület, a nukleinsavak építőeleme. A DNS-ben és az
RNS-ben is megtalálható az adenin (6-amino-purin) és a guanin
(2-amino-6-hidroxi-purin). A nukleinsavakban a pirimidinbázisokkal társulnak
hidrogénkötésekkel: az adenin a citozinnal, a guanin a DNS-ben a timinnel, az
RNS-ben az uracillal.
A purinbázisoknak többféle térszerkezete (laktim,
vegyes és laktám) ismert; ez befolyásolja a tulajdonságukat – nem mindegyike
található meg a DNS-ben/RNS-ben. Ritkán a metilszármazékaik (N6-metil-adenin,
N6-dimetil-adenin, N2-metil-guanin és N7-metil-guanin
stb.) fordulnak elő. A purinbázisok közé tartozik még a xantin
(2,6-dihidroxi-purin) és a hipoxantin (6-dihidroxi-purin) is, de ezek nem
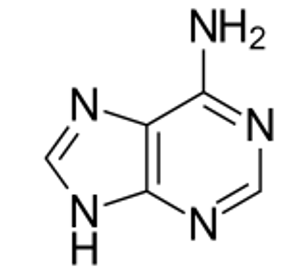
részei a nukleinsavaknak. (→hipoxantin, xantin). Az ábra az adenin szerkezetét
mutatja.

 ▪ A
genetikában a nukleinsavak egyik összetevője: gyűrűs szerkezetű, a szénatomhoz
egyes kötéssel kapcsolódó nitrogéntartalmú szénvegyület. A DNS-ben és az
RNS-ben purinbázisok (adenin [A], guanin [G]) és pirimidinbázisok (citozin [C],
uracil [U] és timin [T]) fordulnak elő. Az emberi DNS ~3 billió bázist
(nukleotidot) tartalmaz.
▪ A
genetikában a nukleinsavak egyik összetevője: gyűrűs szerkezetű, a szénatomhoz
egyes kötéssel kapcsolódó nitrogéntartalmú szénvegyület. A DNS-ben és az
RNS-ben purinbázisok (adenin [A], guanin [G]) és pirimidinbázisok (citozin [C],
uracil [U] és timin [T]) fordulnak elő. Az emberi DNS ~3 billió bázist
(nukleotidot) tartalmaz.