analog hasonmás (→hasonmásság) ■ analogous protein hasonmás fehérje
(→fehérje) ■ analogous gene hasonmás gén
(→gén) ■ analogous/analog organs hasonmás
szervek (→szervek) ■ analogy →hasonmásság.
androgénérzéketlenség androgen insensitivity
syndrome, AIS; testicular feminization X-kromoszómához kötött, lappangó
öröklődésű nemi szervi fejlődési rendellenesség, amelyben a 46 XY kromoszóma szerkezetű férfinek külső női nemi
szervei vannak. (A régi nevét: testicular feminization syndrome,
magyarul herés nőiesedés*, már nem használjuk.) Egyéb jellegzetességek:
▪ A hüvely vakon végződik; lehet csaknem teljes
hosszúságú, általában rövidebb, és egészen csekély is.
▪
Nincs méh.
▪ A belső nemi szervek férfiasak. A herék
szabályosan kifejlődtek, de a hasüregben, ritkán a lágyékcsatornában vannak.
Előfordulnak a nagyajkakban is. Harminc százalékban rosszindulatúan elfajulnak,
feltehetően, mert a szabályos sejtkörnyezetük nem fejlődött ki.
Nagyon ritka betegség (60 000 szülésre esik
egy). Három formáját különböztetik meg: a teljes érzéketlenségűt (complete AIS,
CAIS), a részleges (partial AIS, PAIS) és enyhe érzéketlenségűt (mild AIS,
MAIS).
▪ A teljes androgénérzéketlenségben, amely
messze a leggyakoribb (ha androgénérzéketlenségről beszélünk, általában ezt
értjük rajta), a külső nemi szervek teljesen nőiesek.
▪ A részlegesben valamelyik (a női vagy a
férfi) külső nemi szerv túlsúlyban van, de lehetnek kettősök is.
▪ Az enyhe formában
inkább férfias, de nagyon fejletlen.
Oka
az androgénjelfogó érzéketlensége a jelvivőjével (tesztoszteron,
dihidrotesztoszeron, DHT) szemben. A jelfogó egyáltalán nem, avagy csak
kismértékben köti a jelfogót. Az androgénjelfogót az X-kromoszóma hosszú
karjának 12-es sávjában (Xq12) lévő gén (AR (androgen
receptor) gén) kódolja, melynek gyakori a másulása, ötszáznál is több ismert.
Hetven százalékukat anyai öröklődésűnek tartják, és csak 30% keletkezik
újonnan. Azok okozzák ezt a betegséget, amelyek akadályozzák az androgén
jelfogót, hogy kötődjön a tesztoszteronnal vagy a dihirotesztoszteronnal. Az ép
herék megfelelően képezik a tesztoszteront és a dihidrotesztoszeron, de a
férfiasodás elmarad, mert a jelfogó nem köti meg őket.
A
Müller-cső kifejlődése hiányos, a herék elnyomják: a Sertolli-sejtekben az AMH
(anti Müller-hormon) szabályosan termelődik, és elfojtja Müller-csövek
kialakulását, azok felszívódnak.
A szérum ösztrogénszintje szabályos, hasonló a nők
ösztrogénszintjéhez. Ösztrogén tehát bőven termelődik az androgének ösztrogénné
alakításának következményeként (aromatization). Hatására az emlők kifejlődnek,
a zsírszövet és minden más külső megjelenés női irányú.
Rendszerint serdülőkorban ismerjük fel elsődleges
havibajhiány, görcsök miatti vizsgálatkor. Ritkán gondos újszülött- vagy gyermekkori
vizsgálat már korában fényt deríthet rá. Ennek nagy a jelentősége, főként a
lelki előkészítés miatt. Gyakori a lágyéki sérv, amely a serdülőkor előtt
egyébként ritka – ez felkeltheti a gyanúját.
A
nemi szervek férfiasodása az ébrényi élet 8–14 hetében zajlik, AIS-ben
tökéletlenül.
androgénjelfogó androgen receptor,
AR vagy NR3C4 (nuclear receptor subfamily 3, group C, member 4) sejtmagi jelfogó. A jelvivőit az androgének tevősítik;
bizonyos sejtekben elsősorban a tesztoszteron, például a Wolff-csőben; másokban
inkább dihidrotesztoszteron, DHT, mint a nemi szervi öböl (urogenital sinus),
az ivarredő, ivarsánc, ivargumó és a hajtüsző. Ekként a tesztoszteron felelős
az elsődleges, a dihidrotesztoszteron pedig a másodlagos férfi nemi jelleg
létrejöttért. A jelvivővel a sejtplazmában kötődik, majd a sejtmagba
szállítódik.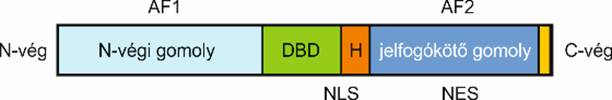
Az androgénjelfogó DNS-kötő átírásfehérje, szabályozza gének
kifejeződését, többek közt a férfi nemi jelleg kialakulását és fenntartását
kialakító génekét. Három fő gomolya van:
▪ Az N-végen az átírást szabályozó, a DNS-kötő és a jelvivőt
kötő gomoly található. Része az AF1 fehérje. A leginkább a N-végfelé lévő igen
változékony.
▪ A DNS-kötő (DBD) törzsökös, jószerivel nem változik, ezért
sajátos feladatot csak a selective androgen response elements (AREs)-szel
kötődve végezhet. Két cinkujj-mintázat van,
ezzel ismer fel törzs bázissorokat az indítóban és a fokozóban.
▪ A C-vég felé a jelvivőket kapcsolók helyezkedek el.
Az X-kromoszóma hosszú karjának 12-es sávjában (Xq12) lévő
gén kódolja, melynek gyakori a másulása, ötszáznál is több ismert. Azok,
amelyek abban akadályozzák az androgén jelfogót, hogy kötődjön a tesztoszteronnal/dihirotesztoszteronnal,
okozzák az →androgénérzéketlenséget.
antigén–ellenanyag
képződmény →immunképződmény
antigénerősség az
antigén fajlagos kötődésének és immunválaszt kiváltó képességének a mértéke.
Erős antigének nevezzük a B- és T-sejtek által is felismerhető antigéneket. Az
antigének erőssége függ:
• Az antigén anyagától, vegyi összetételétől. A
fehérjék a legerősebb antigének (főleg szerkezeti merevségük miatt), a
szénhidrátok gyengébbek (szerkezetük ismétlődő egységekből áll; ezért könnyen
változik), a lipidek nagyon gyenge antigén hatásúak (térszerkezetük nem
rögzült). A DNS-, RNS-részek közül általában a nagyobb molekulatömegű és a
kevéssé metilezett az erősebb antigén hatású, a kisebb tömegűek és a metilezett
formák jóval kevésbé.
• Az antigén vegyi és fizikai (tér-) szerkezetétől, a
szerkezet állandósultságától (rögzült szerkezetek erősebb antigén hatásúak).
• Az antigén nagyságától. A <2000 Da tömegű
molekuláknak kicsi az antigén hatásuk, kivételek, pl. gyógyszermolekulák
azonban vannak.A nagy molekulák viszont erős antigének, mert kevéssé oldódók,
és a falósejtek könnyebben feldolgozzák.
• Az antigének lebonthatóságától (az antigén-bemutató
sejtek csak a lebontható antigénekből képezhetnek kis peptideket).
• Az antigének biológiai eredetétől
(saját, egyedmás*, idegen [auto-, allo-, xeno-] antigének).
antigénfedés* opsonisation (opszonizáció) kórokozók vagy elhalt sejtek antigénjeinek fedése
ellenanyagokkal (IgM, IgG1, IgG3) vagy komplementtöredékekkel (C3b, C3d és C4b)
jelölésre a sejtfaló sejtek számára. Az ellenanyag/komplementtöredék az
antigén-meghatározóhoz (epitope) kötődik, lehetővé téve, hogy kapcsolódjék a
falósejt jelfogójával. Az immunsejtek (falósejtek), az elhalt sejtek és a
kórokozók sejtfelszíne is negatív töltésű, ezért ezek taszítják egymást. Az
antigénfedés ezt a taszítóhatást hidalja át.
antigénfelismerés Csak az immunglobulinok
(ellenanyag, B-sejt-jelfogó [BCL]), a TLR (toll-like receptor), az
MHC-fehérjék, továbbá a T-sejtek, a nagyfalósejtek és a nyúlványos sejtek
ismerik fel az antigént. Az ellenanyagok,
a BCL, a TLR és az MHC közvetlenül azonosítják az antigéneket, közvetlenül
veszik észre az antigén-meghatározókat. A T-sejtek, a nagyfalósejtek és a
nyúlványos sejtek közvetve érzékelik az antigén-meghatározókat.
• A T-sejtek áttételesen, az antigén-bemutató sejtek
közvetítésével azonosítják az antigént; az MHC-vel kapcsolt
antigén-meghatározót ismerik fel.
• A nagy falósejtek nem az antigént észlelik, hanem az
antigén–ellenanyag képződmény Fc-töredékét.
• A nyúlványos sejtek a mannóz-, az Fc- és a DEC205-jelfogóikkal az antigének hordozó részét,
illetőleg az antigén–ellenanyag képződmény Fc-töredékét érzékelik.
A
sejtek közül tehát csak a nyiroksejtek és az antigén-bemutató sejtek fedezik
fel az antigéneket; mindegyik valamelyik felszíni jelfogójával.
antigénhatás Az antigének az
ellenanyagokra és a nyiroksejtekre hatnak. Az ellenanyagokkal kapcsolódva
tevősítik őket, másrészt együtt alkotják az immunképződményeket
(antigén–ellenanyag képződmények). Hatásuk a nyiroksejtekre a nyiroksejtek
érettségétől függ: az éretlen nyiroksejtek érését, az érettek gerjesztését
okozzák; B- és a T-sejtes immunválaszt váltanak ki. (→nyiroksejtérés) Az
antigének a B-sejteket a T-sejtektől függően és tőlük függetlenül is gerjeszthetik.
A T-sejt-függő antigének javarészt fehérjék, serkentő hatásukhoz értelemszerűen
szükségesek a T-sejtek; ezek képezik a gerjesztődés második jelét. A
T-sejt-független antigének szokásosan poliszaharidok, lipidek vagy
nukleinsavak, amelyeket az antigén-bemutató sejtek nem képesek feldolgozni és
bemutatni a T-sejteknek, következésképpen a T-sejtek nem ismerhetik fel őket;
ezért nevezzük ezeket T-sejttől független antigéneknek. Jelölésük TI- (T cell
independent) antigének; két fajtájuk a TI1- és a TI2-antigének. A
kétféle antigénhatás részleteit a B-sejteknél ismertetem.
antigénhatás-erősség Ez a fogalom a nyiroksejtek gerjesztésének és/vagy az
ellenanyag képződésének mértékére vonatkozik. Jelentősége elsősorban az
oltásoknál van; a daganatgyógyászatban a HPV-oltásnál. Az antigénhatás függ:
▪ Az antigénerősségtől és az antigénhatás
időtartamától (minél tartósabb az antigénhatás, annál kifejezettebb).
▪ Az antigén beadás/bejutás módjától: Az antigének más
hatást váltanak ki a nyálkahártyákon és mást a bőrön vagy a bőr alatti
kötőszövetbe jutva. Ugyanaz az antigén (pl. a táplálék fehérjéi) teljesen
eltérő választ válthat ki a bél nyálkahártyáján át bejutva, mint a bőr alá
fecskendezve. Az utóbbinál lassan szívódnak fel, ezért hatásuk kifejezett.
• Az antigén mennyiségétől: az antigén kis mennyiségben
immuntűréshez, túl nagyban az immunrendszer megbénulásához vezethet. Közepes
mennyiségben a legerősebb hatású – a hatáserősség a Gauss-görbe szerinti.
• Genetikai tényezőktől: ugyanazon antigénre adott
immunválasz egyénenként nagyon különbözhet; sőt bizonyos génhibák a T-sejtes
immunválasz elmaradását okozzák (károsodik a nyiroksejtképződés vagy az
antigén-bemutatás).
• Az életkortól: csecsemőknél szokásosan a 6–12.
hónapban alakulnak ki az immunfolyamatok. Időskorban változik az immunológiai
válaszadás.
• Az antigénhatás ismétlődésétől: ez az ún. erősítő
(booster) hatás – az antigén ismételt bevitele (oltásoknál) sokszorosára növeli
hatását a B- és a T-sejtek, valamint az emlékező sejtek kifejezett burjánzása
következtében.
• Oltásoknál az antigén hatékonyságát rendre valamilyen
adalék hozzáadásával serkentik.
antigénszerkezet Az
antigének két részből tevődnek össze: az antigén-meghatározóból és a
hordozórészből.
▪ Antigén-meghatározó (epitope) az antigénnek az ellenanyaghoz és a
nyiroksejtekhez közvetlenül kötődő része, ez felelős az antigén
fajlagosságáért. Az antigén-meghatározó
lehet vonalszerű (linear epitope) vagy térhelyzeti (conformational epitope). A
vonalszerű meghatározó a polipeptid eredeti aminosav sorrendjének 6–8 tagjából
áll; általában a T-sejtek azonosítják. A térhelyzeti meghatározók a fehérjék
negyedleges szerkezetével függnek össze, ugyanis ez teszi lehetővé, hogy távoli
peptidek egymás mellé kerüljenek, és kialakítsanak antigén-meghatározót. A térhelyzeti
meghatározókat szokásosan a B-sejtek észlelik. A szénhidrátokban, a lipidekben,
a nukleinsavrészekben általában elektronkötésű szerkezetek az antigén-
meghatározók. Jószerével a félantigének is antigén-meghatározók. Az
antigén-meghatározók – ellentétben az antigénekkel – mindig az antigén
részeként léteznek, oldott formájuk nincs.
Egy-egy nagy antigén felszínén rendszerint több
antigén-meghatározó van (vonalasak és térhelyzetiek is); ezek mindegyikére
kialakulhat fajlagos, de egymástól független immunválasz. Egyetlen nagy antigén
tehát sokféle immunválaszt válthat ki. Az immunválasz mindig egyetlen
antigén-meghatározóra jön létre és nem a nagy molekulák [pl. fehérje]
antigén-meghatározóinak összességére. A több antigén-meghatározó közül vannak
uralkodó jellegűek, ezek keltik a legerősebb immunválaszt, és háttérbe
szoríthatják a többi antigén-meghatározó által elindított immunfolyamatokat.
▪ Hordozórész az antigén-molekulának az antigén-meghatározón kívüli
része. Részt vesz az immunválaszban.
antigénváltozás az
antigén szerkezetének átalakulása. A kórokozók gyakran
változtatják antigénjeik szerkezetét; ez az egyik formája az immunrendszerrel
szembeni védekezésüknek. A szerkezet változtatásának két, egy lassú és egy
gyors formája ismert.
antigén-eltolódás* antigenic drift a vírusok
antigénjeinek olyan parányi módosulása, mely megváltoztatja az antigén-meghatározó
szerkezetét. Véletlenszerű parányi génváltozások (pontmódosulások) folyamatosak
a vírusokban. Legtöbbször nincs különösebb következményük. Előfordul azonban,
hogy a parányi átalakulás megváltoztatja az antigén-meghatározó fajlagosságát,
és ennek következtében az eredeti antigénnel szemben képződött ellenanyagok már
nem kapcsolódnak hozzá. Ezt nevezzük antigén-eltolódásnak, amely a vírusok
természetes kiválasztódásának eszköze, az immunvédekezés kijátszása érdekében.
antigénváltás* antigenic shift a vírus
antigének átalakulása más antigénekké. Két vagy több vírustörzs, avagy két vagy
több vírusfaj génjeinek átcserélődése, aminek következtében új gén és új
antigén keletkezik; az eredeti antigén-fajlagosság eltűnik. A keletkezett
antigének a szervezet számára teljesen újak; ezért pl. a korábbi oltások már
nem védenek ellene. Az antigénváltás az influenzavírusoknál viszonylag gyakori,
járványos fertőzésekhez vezet. Hasonló folyamatok figyelhetők meg a
rosszindulatú daganatokban is: a kezdetben még erős immunválaszt kiváltó
felszíni antigéneket genetikai módosulások megváltoztatják, ezért a korábban
keletkezett ellenanyagok már nem kötődnek hozzá.
belrokonmásság*LM paralogy génkettőződés (duplication event) következtében kialakult
rokonmásság.
belrokonmás*LM paralog/paralogous belrokonmásságban lévő bázissor, gén, fehérje, RNS.
belrokonmás bázissorok*LM paralogous
sequences valamely fajban génkettőződésből keletkező, többé-kevésbé hasonló
bázissorok. (→rokonmásság)
belrokonmás gének*LM paralogous genes (gene paralogs) olyan
többé-kevésbé hasonló gének, amelyek kettőződésből jöttek létre; szerkezetük
és/vagy tevékenységük változott. A gén kettőződik, hogy a DNS-ben két helyet
foglaljon el. A génkettőződés valamely fajban keletkezik, szokásosan
szükségszerűségből, de belrokonmás gén előfordulhat két különböző fajban is, ha
a fajkeletkezés a kettőződés után ment végbe. (→rokonmásság)
belrokonmás fehérjék*LM paralogous
proteins (protein paralogs) belrokonmás gének által kódolt fehérjék. (→gén)
BRCA gének BRCA genes a BRCA1 és a BRCA2 gén. A
BRCA1 a 17-es kromoszóma hosszúkarján (17q21), a BRCA2 a 13-as kromoszóma
hosszúkarján található (13q12-13) daganatgátló gén. Mindkettő nagy,
~80 000 bázispárból áll. A kódoló szakaszok nagysága 5,6 (BRCA1), illetve
10,2 kb (BRCA2). A BRCA1 gén 22 képezőt* (exont) tartalmaz, nyolc változata
van; az 1-es, 2-es változatának mérete 1863 aminosav. A BRCA2 gén 27 képezőt
tartalmaz, és 3418 aminosavból álló fehérjét képez. Ezeknek meghatározó
szerepük van a kétszálú DNS-törés hasonmás átrendeződéses* kijavításában. A
BRCA1 a helyreállítás kezdetén, a javítófehérjék vonzásában és rögzítésében,
továbbá a sejtkör szabályozásában, a BRCA2 pedig a későbbi szakaszában, a
DNS-szál tört végeinek egyesítésében vesz részt. Szerepük
van a génátíródás szabályozásában is, valamint a kromatin módosításban A BRCA1
az 53BPI (TP53 binding protein) fékezésével elősegíti, hogy a sejt a kétszálú
DNS-törést a hasonmás átrendeződéssel javítsa ki.
coding region microsatellites, cMS génbeli
mikroismétlet (→mikroismétlet)
consensus, másképpen canonical, a molekulabiológiában törzs consensus gene törzsgén
(→gén) consensus
protein törzsfehérje (→fehérje) consensus sequence (→törzssor) consensus splicing site
(→RNS)
degeneracy of genetic code →génkódbőség
discontinuous,
split genes közteses gének* (→gén)
egygénes öröklődés egyetlen gén által meghatározott jelleg átadása az
utódnak. Négyféle ismert:
▪ Uralkodó öröklődés (dominant
inheritance) (domináns öröklődés) uralkodó válzat által közvetített
genetikai jelleg továbbítása a magzatba. A válzat párja nem befolyásolja a
megnyilvánulását; ha az uralkodó válzat jelen van, mindig kifejeződik. Az
uralkodó válzat származhat az anyától vagy az apától, és 50%-os valószínűséggel
adódik tovább; így betegséget okozó másult uralkodó válzat, ha átadódik, az
utódban is mindenképpen kiváltja a betegséget. Példa erre a Huntington-kór,
amely tehát akkor is kialakul, ha a másik szülőtől az ép válzat öröklődik, azaz
beteg szülő esetében 50% valószínűséggel lesz az utód is Huntington-kóros.
Mivel ezek a betegek a természetes kiválasztódásban eltűnnek, az uralkodó
öröklődésű betegségek jóval ritkábbak, mint a lappangó öröklődésűek.
▪ Lappangó öröklődés (recessive
inheritance) (recesszív öröklődés) lappangó válzat által továbbított
genetikai jelleg, amely csak akkor nyilvánul meg, ha a válzat párja is
lappangó, azaz párban van jelen; vagy ha az uralkodó válzatpárja elvész
(válzatvesztés) vagy elnémul, például módosulás, másulás vagy gátlóhatások következtében.
▪ Kifejeződő öröklődés* (codominant
inheritance) két együttesen kifejeződő génválzatpár hozza létre. Mindkét válzat
(szemben a köztes öröklődéssel) ugyanolyan mértékben kifejeződik, mintha
külön-külön határoznák meg a jelleget. Például az AB-vércsoport: az
AB-vércsoportú egyénben a vércsoportot meghatározó gén két válzata, egy A- és
egy B-válzata van jelen. A kettősük által meghatározott vércsoport esetében
(például vérátömlesztésnél) egyforma mértékben (és nem köztes állapotként)
nyilvánulnak meg mind az A, mind a B vércsoport sajátságai, mindkettőt
figyelembe kell venni, azaz egy harmadikféle (AB-vércsoport) változat
keletkezik.
▪ Köztöröklődés (intermediate inheritance, incomplete dominant
inheritance) egyenlően kifejeződő válzatok hozzák létre, egyik sem nyomja el a
másikat, a keletkező jelleg a kettő közötti. Ez úgy képzelhető el, hogy a
génpár egyike által képzett fehérje szintje sem 100%-os; a két fehérje
50–50%-ban van jelen, és mindegyik érezteti hatását.
▪ Többválzatos öröklődés (multiple allele inheritance) a génnek az adott fajban
kettőnél több válzata létezik; az egyed genetikai jellege attól függ, hogy
melyik két válzatot örökölte. Szintén példája az AB0 vércsoport. A három
leggyakoribb válzat A, B és 0; ebből kettő található meg egyetlen egyedben, és
ez a kettő határozza meg a vércsoportot: Ha A és A illetve A és 0 válzat van
jelen, a vércsoport A (az A válzat az uralkodó a 0-val szemben). Ugyanígy, ha B
és B, illetve B és 0 válzat van jelen, a vércsoport B (a B válzat szintén
uralkodó a 0-val szemben); ha két 0 válzat van, a vércsoport 0; míg ha A és B
van együtt, a vércsoport AB (l. fent).
ellenanyag–antigén kapcsolódás az
ellenanyag Fab-szakaszának kapcsolódása az antigénhez a változó (VH
és VL) gomolyokkal. (→ellenanyagok szerkezete) A VH és VL
gomolyokban vannak állandó sorrendű aminosavszakaszok (framework region [FR])
és három változó sorrendű szakasz (complementarity-determining regions [CDRs];
CDR1, CDR2 és CDR3); a CDR3 a legváltozékonyabb. A VH és VL
hat változó aminosavsora együtt ismeri fel és köti meg az antigén-meghatározót,
mindegyik karon külön. Vagyis egy ellenanyag – mivel két Fab-szakasza van – két
azonos vagy különböző antigén-meghatározóhoz kapcsolódhat, jelentősen fokozva
az antigének hatékonyságát. Kapcsolódhat pl. egy kórokozó két antigénjéhez,
vagy két kórokozóhoz.
A hat változószakasz háromirányú „zsebet” (three-dimensional
pocket) alkot; az antigén ebbe illeszkedik saját tükörképeként. A zsebek
mintázódnak fajlagosan az antigén-meghatározóknak megfelelően, azaz ezek
felelősek az ellenanyagok fajlagosságáért. A társult ellenanyagok, mint pl.
sIgA, sIgM, több ponton is kötődhetnek. A kapcsolódás visszafordítható, gyenge
kötésekkel (hidrogénkötések, töltésvonzás, sóhidak, van der Waals-kölcsönhatás)
jön létre.
A kötődések mértékét kétféle mutatóval jellemezzük:
• Kötéserősség* (affinity, affinitás) egyetlen
Fab és az antigén kötődésének erősségét, vagyis az egy pontban (monovalent) kötődés
erősségét fejezi ki. Az antigénhez erősen kapcsolódó ellenanyagok (high
affinity antibodies, antigénfogékony ellenanyagok) gyorsan hatnak, és már
kevesebb is hatásosabb a fertőzéssel szemben, mint a gyengébben kötődőkből (low
affinity antibodies).
• Összkötéserősség* (avidity, aviditas) az ellenanyagok
összessége által létrehozott kapcsolódás erőssége, vagyis a több pontban
(multivalent) kapcsolódás összesített erőssége. Függ a kötéserősségtől, a
kapcsolódások számától és az antigén, ellenanyag szerkezetétől.
Megkülönböztetünk elsődleges és másodlagos
antigén–ellenanyag kölcsönhatást: az elsődleges az antigén és az ellenanyag
kötődésének közvetlen következménye, például az ellenanyag kapcsolásakor
megváltozik az antigén szerkezete. A másodlagos az antigénhez kapcsolódott
ellenanyagok által kiváltott hatás, például az antigénhez kötődött
immunglobulinok Fc-részeinek végrehajtó sejteket gerjesztő hatása.
ellenanyag–antigén képződmény →immunképződmény
epitope antigénmeghatározó (→antigén)
fajrokonmásság orthology
a törzsfejlődésben fajelkülönülés (speciation event, orthologs) következtében
létrejövő rokonmásság. Olyan többé-kevésbé hasonló bázissor vagy az általa
kódolt fehérje/RNS, amelyik közvetlen elődtől adódik át a szétváló fajokba.
A fajrokonmás fehérjéket, géneket stb. angolul conserved-nek
is nevezik; magyarul törzsökös*, pl. törzsökös fehérje/gén.
(→törzsökös)
fajrokonmás ortholog/orthologous fajrokonmásságban
lévő bázissor, gén, fehérje, RNS.
fajrokonmás bázissor orthologous sequences fajelkülönülésben (speciation
event, orthologs) az elődfajból átadódó bázissor; fajelkülönülésből származó
rokonmás bázissor. A fajrokonmás bázissor a két fajban eltérővé válhat, vagyis
nem minden részletében ugyanaz. (→bázissor)
fajrokonmás gének* orthologous genes (gene orthologs) különböző
fajokban lévő, közös őstől származó gének; fajelkülönülésből (speciation event,
orthologs) származó rokonmás gének. Ezek szerkezet és az általuk kódolt
fehérje/RNS szempontjából is viszonylag hasonlóak, de lehetnek lényegesen eltérőek
is. (→gén)
fajrokonmás fehérjék* orthologous
proteins (protein orthologs) a törzsfejlődésben fajelkülönülés (speciation
event, orthologs) következtében elkülönült, közös elődtől származó gének által
kódolt fehérjék; fajelkülönülés következtében létrejövő rokonmás fehérjék. Az
ilyen fehérjék aminosavlánca hasonló, legalábbis közös jellegzetes
aminosavmaradék-soruk van. A fajrokon
fehérjék működése – szerkezeti hasonlóságuk ellenére – eltérhet. (→fehérje)
founding mutation alapító génhiba (→génhibák)
főszövetegyező géncsoport (egyéb
nevei: MHC gének, HLA) major histocompatibility
complex, MHC a 6-os kromoszóma rövid karján (6p21) elhelyezkedő géncsoport,
amely olyan sejtfelszíni fehérjéket kódol (MHC-fehérjék), amelyek segítik az
immunrendszert a testidegen molekulák felismerésében; a testidegen és a
testazonos molekulák megkülönböztetésében. A géncsoport 4 Mb hosszú (a DNS
0,13%-át teszi ki), igen sok gént foglal magában, mindegyiknek számos válzata
ismert. Ez teszi lehetővé az MHC-fehérjék sokféleségét, az egyének közti
különbözőségét. Az emberi MHC géneket HLA-géneknek (human leukocyte
antigens, HLAs)
nevezzük. Két osztályukat, az MHC1- és az MHC2-osztályt (class I és class II
MHC complex) különítik el.(→MHC-fehérjék)
hasonmásság analogy a biológiában nem
közös elődtől/őstől származó két vagy több hasonló szerv (pl. a madár és a
darázs szárnya: mindkettő azonos feladatra szerveződött, de nem közös eredetű),
molekula vagy azok részének viszonya. Az orvostudományban a hasonmásság
fogalmát leginkább a nem közös eredetű, de többé-kevésbé hasonló feladatú
és/vagy szerkezetű molekulákra vonatkoztatjuk. Általában környezeti hatásra, a
környezethez való hatékonyabb alkalmazkodás érdekében, de egymástól függetlenül
jöttek létre (convergent evolution).
hasonmás* (biológiai) analog
olyan nem közös eredetű szerv, molekula stb., amely szerkezetében és/vagy
tevékenységében igen hasonló. Közös feladatra szerveződött.
hasonmás fehérjék* analogous
proteins (protein analogs) szerkezetében
és vagy működésében hasonló, de más-más eredetű fehérjék. Ezeket nem közös
eredetű gének kódolják. (→fehérje)
hasonmás
gének* analogous genes (gene analogs) hasonló
feladatú fehérjét kódoló, de nem közös őstől származó (nem azonos géncsaládba
tartozó) gének. (→gén)
hasonmás
RNS* analogous RNA kizárólag RNS örökítőanyagú
vírusokban fordul elő. Nem közös őstől származó, hasonló tevékenységű RNS-ek.
hasonmás
szervek*
analogous/analog organs ugyanarra a feladatra szerveződött, de nem közös
eredetű szervek, pl. a madarak és a méhek szárnya.
hasonmás
vegyület* analog, analogous compound olyan vegyület,
amely szerkezetében hasonlít egy másikhoz, de összetevőiben egy-két ponton
különbözik: egy vagy több atom/hatócsoport más. Pl. hasonmás gyógyszerek
(analog drugs): szerkezetükben hasonló, de vegyi és biológiai tulajdonságukban
eltérnek. Ismerünk nukleotid hasonmásokat, ezek kicsit mások, mint a szokványos
bázissorú nukleotidok, de képesek beépülni a DNS megfelelő részébe, viszont más
kölcsönhatásokat létesítenek.
hidrogénkötés (hidrogénhíd, H-híd) összekapcsolás
hidrogénatom részvételével: az egyik molekula hidrogénje kapcsolódik a másik
molekula nagy elektronegativitású atomjának szabad elektronpárjához (az
elektronkötésben nem résztvevő vegyérték elektronpár); hidat képez közöttük.
Viszonylag erős (~0,2 eV, 12–30 kJ/mol) kapcsolódás: egy nagyságrenddel haladja
meg a kétsarkú kölcsönhatások erejét, és egy nagyságrenddel gyengébb az
elektronkötés erejénél.
Oka az, hogy a
hidrogén protonja meglehetősen kicsi, ennek következtében a pozitív töltése
kifejezett, hiszen a töltés jóval kisebb felületen oszlik el, mint a nagy
protonoknál. Az elektronkötésben a hidrogén az egyetlen elektronjával vesz
részt, protonja viszonylag szabaddá válik, és erős vonzerejével vonzza a
közelében lévő elektronegatív atom elektronjait, lehetővé téve a kapcsolódást.
A hidrogénkötés kialakulásának tehát három feltétele van:
▪ olyan molekula,
amelynek valamelyik elektronkötésében hidrogén vesz részt,
▪olyan másik
molekula, amelynek van nem kötő elektronpárú, erősen elektronegatív atomja,
▪és hogy a két
molekula viszonylag közel (0,3 nm) kerüljön egymáshoz.
A hidrogénhíd csak az oxigén, a nitrogén
és a fluor (pillératomok) között létesül – a feltételek csak ezek
kapcsolódásaiban adottak. A hidrogénkötéshez szükséges részpozitív hidrogén
jóformán csak az O–H, N–H, F–H kötődésekben keletkezik, a hidrogénkötés pedig
csakis az O–H, N–H és F–H csoportok O–H, N–H és F–H csoportokkal való
társulásakor jöhet létre: pl.
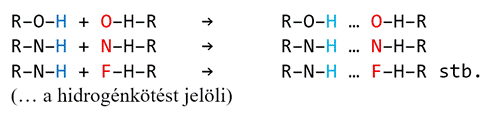
Más, például a szénatomhoz (C) kapcsolt
hidrogén nem képes hidrogénkötésre, mert a C–H csoport elektromosan semleges:
az elektronok eloszlása az atomok között egyenlő. Jóllehet részleges
pozitivitás, negativitás ezekben is létrejöhet, de ez nem elegendő a
hidrogénhíd kialakításához.
A hidrogénhíd
általában molekulák között jön létre, pl. DNS-ben, fehérjékben, de előfordul
molekulán belül is, pl. az acetilacetonban.
hidrogén-peroxid hydrogen peroxide gyenge sav, erős
elektronelvonó; szerepe van a szabadgyökök keletkezésében. Képlete: H2O2. A víznél sűrűbb, halványkék,
oldatban szagtalan, színtelen és keserű ízű, párolog (párolgása a szemeket
ingerli). Fertőtlenítő hatású, az orvosi gyakorlatban pl. a sebek
fertőtlenítésére használjuk.
homo- ■ homogeneous egynemű ■ homologous (homolog) rokonmás ■ homologous chromosome kromoszómapártag
(→kromoszóma) ■ homologous protein rokonmás
fehérje (→fehérje) ■ homologous gene rokonmás
gén (→gén) ■ homologous recombination →szülőmás átrendeződés* ■ homologous recombination repair
(HRR) →rokonmás átrendezés* ■ homologous
RNA rokonmás RNS (→RNS) ■ homologous
sequence rokonmás bázissor (→bázissor) ■ homology →rokonmásság
Hoogsteen-féle hidrogénkötések Hoogsteen hydrogen bonds a DNS-ben előforduló olyan hidrogénhidak, amelyek
mások, mint a DNS szálait összekötő, vagyis az adenin és a timin, illetve a
guanin és a citozin között létrejövő hidrogénkötések (Watson–Crick-féle
hidrogénkötések). Ilyenek alakulhatnak ki, pl. ha a purinbázis elfordul, és az
5-ös gyűrűje kapcsolódik pirimidinhez, nem pedig a hatos gyűrű, avagy két
azonos purinbázis (adenin–adenin; guanin–guanin) között stb. (→G-négyes)
A Hoogsteen-féle
hidrogénkötések másodlagos DNS-szerkezeteket hoznak létre a DNS számos helyén.
Ezek többnyire ingatag szerkezetek, sokszor csak átmenetiek, de a hozzájuk
kapcsolódó fehérjék rögzíthetik, állandósíthatják.
Jelentősen megváltoztathatják
a bázispár viszonyait is, ami befolyásolhatja a fehérjék általi felismerésüket,
a DNS-hibajavítást, a DNS-kettőződést. (→DNS-szerkezet)
housekeeping gene háztartási gén* (→gén)
inzulin/IGF1 és az androgének Kétféleképpen befolyásolja az androgének képződését. Egyrészt a
GnRH–LH/ACTH vonalon serkenti, másrészt közvetlenül az inzulinjelvonalon, amely
– mint a fentiekben ismertettem – serkentő és gátló is lehet.
• Növeli a tüszők toksejtjeiben az androgének
képződését; fokozza a szteroidképző sejtekben a CYP17A1 és a 3βHSD enzimek működését. A CYP17A1 hatása az LH-hoz
kapcsolódva hatványozódik, amelynek magyarázata az inzulin és az LH
jelátvitelének kapcsolódásában keresendő.
• GTH-szerűen, leginkább LH-ként hatva serkenti a
tüszők tok- és granulozasejtjeit; ennek is szerepe van az androgénképzés
fokozásában.
• Fokozza az agyalapi sejtek érzékenységét a GnRH-val
szemben, és megzavarja a szabályos GTH-kiáramlást.
• Növeli az ACTH hatását az androgénképző sejtekre:
ACTH adásakor a mellékvesekéreg sejttenyészetében több androgén képződik, ha
inzulint is adnak.
▪ Az inzulintöbblet csökkenti az SHBG keletkezését,
ezáltal lesz több szabad tesztoszteron, amelynek szerepe van a kórkép
kialakulásban.
• A CRH–ACTH rendszert nem módosítja.
közteses gének* discontinuous, split
genes (→gén)
nested gene beágyazódott
génSP (→gén)
nitrogén-monoxid nitric
oxide vízben kevéssé, zsírban jól oldódó gáz. Párosítatlan elektronja van
(szabad gyök), ezért életideje rendkívül rövid (<30 másodperc). A sejtfal
lipidrétegén könnyen átjut, a sejtek közt szabadon terjed. Sok sejtfolyamat
szabályozója, a sejtben az guanilát-ciklázhoz kötődik. A kapcsolódás hatására a
GTP-ből cGMP keletkezik, amely számos jelközvetítés folyamatosítója. (→cGMP, guanilát)
oncogene (onkogén) daganatserkentő gén (→gén)
ortholog fajrokonmás (→fajrokonmásság) ■
orthologous genes fajrokonmás gének (→gén) ■ orthologous proteins (protein orthologs) fajrokonmás fehérjék (→fehérje) ■ orthologous sequences fajrokonmás bázissorok
(→bázissor) ■ orthology fajrokonmásság
örökletes
daganatokat vizsgáló sokgénes készletek* multigene hereditary cancer testing panels Gyárilag összeállított olyan készletek, amelyek
számos, az öröklődő daganatokban szerepet játszó gén egyidejű vizsgálatát
teszik lehetővé. Módszertanilag NGS alapúak. Például örökletes emlő- és
petefészekrák esetén a BRCA1 és BRCA2 gén mellett további 23 gén, továbbá egy
génszabályozó rész vizsgálatára alkalmas (→PALB2, CHEK2, BARD1, BRIP1, RAD51C, RAD51D, TP53, MRE11A, RAD50, NBN, FAM175A, ATM, STK11, MEN1, PTEN, CDH1, MUTYH, BLM, XRCC2, MLH1, MSH6, PMS2, MSH2, 3’ UTR of EPCAM).
ösztrogéntöbblet az
ellensúlyozatlan ösztrogénhatás és az összösztrogénhatás fogalmának közös
elnevezése.
ellensúlyozatlan ösztrogénhatás unopposed estrogen az ösztrogén–progeszteron arány eltolódása az ösztrogén javára. A
megfelelő ösztrogén–progeszteron arány a női nemi szervek szabályos működésének
letéteményese; eltolódása zavart kelt. Három formáját különböztetjük meg:
▪ Rövid idejű erős ösztrogénhatás. Veszteglőtüsző
következménye. A többé-kevésbé szabályosan érett tüsző (Graaf-tüsző) nem reped
meg, tovább növekszik (veszteglőtüsző, persistent follicle), és bőséggel képez
ösztrogént. Ennek hatására, viszonylag rövid idő alatt a méhnyálkahártya
gyorsan nő, jelentősen megvastagszik, túlteng (endometrial hyperplasia).
▪ Tartós gyenge ösztrogénhatás. A tüszőérés zavarának
következménye. A tüszők nem érnek meg teljesen, idő előtt visszafejlődnek,
ezért nem keletkezik sárgatest. Ilyenkor egyszerre több tüsző érhet különböző
mértékben, vagy egyik érését, pusztulását egy másik érése, majd sorvadása
követi, és ezek legkülönbözőbb változatai fordulhatnak elő. A tüszők nem
termelnek egyszerre sok ösztrogént, sőt kevesebbet, mint a szabályosan érők, de
mivel az elváltozás hetekig, hónapokig, sőt éveken keresztül is tarthat, az ösztrogénhatás
összeadódik a méhnyálkahártyában.
▪ Kimerült
petefészeki ösztrogénhatás*. A változókorban a vérzés megszűnése a petefészkek
kimerülésének a következménye: megszűnik a tüszőérés, a tüszőrepedés, a sárgatest
képződése. A petefészkek ösztrogénképzése azonban nem áll le; kevés ösztrogén
az érésnek induló, de gyorsan elsorvadó tüszőkben még évekig képződhet,
többé-kevésbé folyamatosan. Ennek hatására a méhnyálkahártya valamelyest
burjánzik, rendszerint szabálytalanná válik. A csekély mértékben növekvő
méhnyálkahártya fenntartására elegendő a képződő ösztrogén, és mert lényeges
ösztrogéningadozások sincsenek, a méhnyálkahártya nem válik le, nem keletkezik
vérzés. A tüszőállomány teljes kimerülésével az ösztrogénképződés is megszűnik,
és a méhnyálkahártya visszafejlődik, elsorvad.
összösztrogénhatás (élethossznyi
ösztrogénhatás) lifetime estrogen
exposure a nő élete folyamán képződött, méhnyálkahártyát érő összes ösztrogénhatás.
A hóvérzérzések* számával és az
állapotossággal függ össze. A korai serdüléssel és/vagy késői
vérzéselmaradással többször van havibaj, összességében sok ösztrogén hat a
méhnyálkahártyára. A várandósság alatt nincs méhnyálkahártya, ezért a szülések
számával lényegesen kevesebb a méhnyálkahártyát érő összes ösztrogénhatás.
paralog belrokonmás*LM ■ paralogous
genes belrokonmás gének (→gén) ■ paralogous
proteins (protein paralogs) belrokonmás fehérjék (→fehérje) ■ paralogous sequences belrokonmás bázissorok
(→bázissor)
párhibajavító gének*
mismatch genes a párhibajavításban résztvevő fehérjéket (MSH2, MSH3, MSH6, MLH1, MLH3, PMS2 stb.) kódoló
gének. Sokféle van, sokféle kromoszómán. A nevezet nemcsak a párhibajavító
fehérjéket kódoló géneket foglalja magában, hanem azokat is, amelyek a társuló
fehérjéket képezik.
PAX (
paired bo
x) gének Pax genes átírásfehérjéket kódoló géncsalád.
Jellegzetességük a párdoboz* (paired box), valamint a párszerű
hasondoboz* (pair-typed homeobox) és a nyolcpeptidkódoló bázissor (octapeptide-coding
region). Az utóbbi kettő megléte szerint négy alcsaládot különböztetünk meg:
▪ az elsőbe a PAX1 és PAX9 tartozik – nincs bennük
párszerű hasondoboz;
▪
a másodikat a PAX2, PAX5, PAX8 alkotja – a párszerű hasondoboz csak részben van
meg;
▪
a harmadikban a PAX3 és PAX7 van – tartalmazzák mind a három elemet;
▪
a negyedikbe pedig a PAX4-et és a PAX6-ot soroljuk – belőlük a
nyolcpeptidkódoló bázissor hiányzik.
Összesen
tehát 9 PAX gén ismert, ezek nem csoportosan helyezkednek el: PAX5 9p13.
Működésük az ébrényi fejlődés lezárulásával többnyire befejeződik, néhány
szövetféleségben azonban megmarad. A PAX gének másulása, működészavara
összefügg betegségek, pl. rákok kialakulásával.
polygenetic
inheritance, quantitative inheritance →
többgénes öröklődés
pseudogenes álgének (→gén)
rákgéntár*
The Cancer Genome Atlas (TCGA) a National Institutes of Health, U.S. Department
of Health and Human Services két részlege, a National Cancer Institute (NCI) és
a National Human Genome Research Institute (NHGRI) által összeállított olyan
adattár, amely a rákokban előforduló géneltéréseket tartalmazza. A rákgenetikai
kutatás és a rákgenetika gyakorlati alkalmazhatóságának tanulmányozására hozták
létra. Ez a hatalmas (~3 petabite) adattár a kutatók számára szabadon
hozzáférhető.
rendszer, környezet system, surroundings természettudományi fogalmak. A rendszer a világ (az
anyagi valóság) vélt vagy valós határfelülettel elkülönített része, amelyet
magunk választunk ki. Ilyen értelemben számtalan rendszer lehet, fizikai, vegyi
és biológiai rendszerek sokasága. Például kémcsőben lévő folyadék, terem és
tartozékai, vagy akár a Föld légköre, avagy gáz a léggömbben rendszer (a gáz
eloszlásának, mozgásának és a léggömb falának a rendszere). Ami a rendszeren
kívül van, az a környezet (surroundings). Vegyi rendszer például a vegyi
anyagok keveréke, környezete az edényzet és minden más körülötte. Biológiai
rendszer például a nyirokkeringés, az immunsejtek rendszere, környezete a
szervezet többi része. Elemi biológiai rendszer a sejt; környezete a sejtkörüli
állomány.
A rendszert állapotjellemzőkkel írjuk le, mint
hőmérséklet (T), nyomás (p), térfogat (V), anyagmennyiség (n) stb. Ennek
alapján lehet állandó nyomású (izobar), állandó hőmérsékletű (izoterm)
és állandó térfogatú (izochor).
A rendszer és a környezet kölcsönhatása szerint
háromféle rendszert különböztetünk meg: elszigetelt, zárt és nyílt rendszert.
▪ Elszigetelt a rendszer, ha a határfelületén sem
anyag, sem energia nem léphet át. Tehát a rendszerrel energiát sem közölhetünk,
és nem is vonhatunk el.
▪ A zárt rendszer határfelületén energia átléphet, de
anyag nem. Tehát energiát cserélhet a környezetével, például melegítjük. A zárt
rendszer lehet állandó és változó térfogatú. Állandó például egy lezárt kémcső;
térfogata nem változik. A dugattyús henger térfogata változik, függően a
dugattyú állásától.
▪ A nyílt rendszer határfelületén az anyag is átléphet,
nem csak az energia. Ilyen például egy folyadékot tartalmazó nyitott kémcső.
Adhatunk hozzá vagy kiönthetünk belőle folyadékot. Melegítéssel pedig
energiaváltozást idézünk elő.
A biológiai rendszerek nyílt rendszerek.
A
kölcsönhatást a rendszer fala határozza meg, például a hő szempontjából a fal
lehet hőszigetelő (adiabatikus) – ilyen az elszigetelt rendszer fala;
féligáteresztő (szemipermeábilis) vagy éppen hővezető.
A
rendszereket csoportosíthatjuk a rendszer mérhető tulajdonságainak térbeli
eloszlása szerint is:
▪ Egynemű rendszer homogeneous system (homogén
rendszer): mérhető tulajdonságai (állapotjellemzői) minden ponton
egyformák. Például egy jól elegyített oldat – hőmérséklete, nyomása stb. minden
részében azonos.
▪ Egyenetlen rendszer nonhomogeneous system (inhomogén
rendszer): mérhető tulajdonságai folyamatoson, lépésről lépésre változnak.
Például ha egy fémdarabot melegítünk, annak hőmérséklete folyamatosan nő, nem
ugrásszerűen változik.
▪ Egyveleg rendszer heterogeneous system (heterogén
rendszer): mérhető tulajdonságai ugrásszerűen változnak. Például olvadó
jég, amely az olvadása egy pontján folyadékká (víz) alakul, állapota
ugrásszerűen változik. Egyveleg, mert szilárd és folyékony összetevői is
vannak.
rendszerállapot* system state, thermodinamic state a rendszer mérhető
fizikai tulajdonságainak összessége egy adott pillanatban. Függetlenül attól,
hogy az állapot miként alakult ki.
rendszeregyensúly* system equilibrium a rendszert alkotó
részecskék egyenletes eloszlása. Az állapothatározók egyike sem változik.
szénhidrogének hydrocarbons csak szén- és hidrogénatomokból álló szerves
vegyületek, amelyek összekapcsolódva láncokat képeznek (nyíltláncú ~) vagy
gyűrűs szerkezeteket (zártláncú ~) alakítanak ki. Megkülönböztetünk telített és
telítetlen szénhidrogéneket (saturated, unsaturated hydrocarbons). A telített
szénhidrogénekben csak egyes kötések vannak, nincs szabad kötés. A
telítetleneknél szabad kötéshelyű szénatom van, egy hidrogénatom hiányzik. A
telített nyíltláncú szénhidrogének az alkánok, a telített gyűrűs szénhidrogének
a cikloalkánok (gyűrűs alkánok). A telítetlen nyíltláncú szénhidrogének az
alkének és alkinek, a telítetlen gyűrűs szénhidrogének az alkánok.
többgénes öröklődés polygenetic inheritance, quantitative inheritance
olyan jelleg öröklődése, amelyet két vagy több gén határoz meg. Mindegyik
kifejeződik, az általuk kódolt fehérjék együtt hozzák létre az egyedségi
jelleget, például a testmagasságot, vérnyomást, értelmi képességet; ezekben
számos gén által kódolt fehérje vesz részt, a hatásuk összeadódik. A
keletkezett jellegnek számos változata lehetséges; a változatok folyamatosak,
és számszerűsíthetők, például a magasságot cm-ben fejezzük ki.
törzsökös*
biológiai értelmezésben a törzsfejlődésben a fajokban tartósodott, jószerivel változatlan maradt.
törzsökös gén/bázissor* conserved, highly conserved gene/sequence (→bázissor, gén)
trans- ’át, keresztül’ jelentésű előtag ■ trans-acting
regulatory elements, TREs bázissor-szabályozók
(→bázissor-szabályozás) ■ transcript →átirat ■ transcription transzkripció →átírás ■
transcription factor transzkripciós
faktor →átírásfehérje ■ transcription
start site, TSS átíráskezdethely* (→indító) ■ translation, transzláció →átfordítás
■ translocation transzlokáció áthelyeződés
(reciprocal translocation átcserélődés*) ■ translocon hártyajárat* (→plazmahálózat) ■ transmembrane
(protein) hártyaátjáró (→fehérje) ■ transposon,
transposable elements (transzpozon) →ugrálat
vegyülő oxigénelemek*
reactive oxygen species, ROS oxigént
(O2-t) tartalmazó rendkívül vegyülő atomok, molekulák, ionok,
gyökök. Ilyenek a szuperoxid-anion (∙O2-), a
peroxidion (O2-2), a hidrogénperoxid
(H2O2), a hidroxilgyök (∙OH), a hidroxidion (OH-),
a nitrogénoxid (NO∙), maga az oxigénmolekula is stb. (A ∙ a párosítatlan elektront
jelöli.) Közülük a párosítatlan elektront tartalmazók a szabadgyökök;
sejtbiológiai szempontból a
szuperoxid-anion (∙O2-) és a hidroxilgyök (∙OH) jelentős.
A nemzetközi reactive
oxygen species, ROS fogalmán a
gyakorlatban az oxigént tartalmazó szabadgyököket értik (oxygen radicals),
jelentőségüket ebben az értelemben tárgyalják, de a nevezet magában foglalja a
páros elektronú vegyülő oxigénelemeket is. A vegyülő oxigénelemek
nevezetet is e kettős jelentésben értelmezzük.
A vegyülő oxigénelemek az oxigén-anyagcseréből származnak, leginkább melléktermékeként szuperoxid formájában. A többi a
szuperoxidból keletkezik. Pl. ∙O2- és H2O2
keletkezhet a xantin-oxidáz, az aminosav-oxidázok stb által sarkallt
folyamatokban. Az oxigénmolekula vízzé alakulásához 4 elektronszállítóval
továbbított elektron szükséges. Ez többlépcsős folyamat. (→befejező elektronleadás) Az oxigén elektronfelvltele azonban nem mindig megy végbe
tökéletesen: az oxigén részlegesen vesz fel elektront, a hiány megszüntetésére
azonnal vegyül a környezetében lévő molekulával. Előfordul az is, hogy pl. az
energiatermecsi légzési láncből szabadul el elektron a sejtplazmába, ahol
oxigénnel egyesül. Mindkét esetben vegyülő oxigénelemek (szabadgyökök)
keletkeznek. A vegyülő oxigénelemek tehát enzimfolyamatokkal és azok nélkül is
keletkezhetnek.
A vegyülő oxigénelemek a sejtekben alacsony szinten
vannak jelen. Az egyensúlyt az
energiatermecsben, a sejtplazmában és a sejten kívül is megtalálható szuperoxid-diszmutáz
teremti meg elektron leadás-felvétel folyamatban (superoxide
dismutase-catalyzed reaction); az energiatermecsi folyamathoz hasonlóan a
feleslegben képződő szuperoxid-aniont oxigénné és peroxiddá alakítja: 2 O2-
+ 2 H+ → O2 + H2O2. A
hidrogénperoxidokat a katalázok kétlépéses folyamattal alakítják vízzé és
oxigénmolekulává (2 H2O2 → 2 H2O + O2).
A vegyülő oxigénelemeknek szerepük van többféle
jelzésfolyamatban, pl. a sejtvégzet, a génkifejeződés irányításában, de jelvivő
molekulák is lehetnek.
Túlzott felszaporodásuk a sejtekben, pl.
sejtártmányban, energiatermecsi zavarokban, súlyos károsodást, betegségeket
okoz. Serkenthetik vagy gátolhatják a rákképződést, függően a sejtbeli
helyüktől és mennyiségüktől. (→szabadgyökök)
